ديناصور
| الديناصورات | |||
|---|---|---|---|
| التصنيف العلمي | |||
| أصنوفة غير معروفة (أصلحها): | الحياة | ||
| مملكة: | الحيوانية | ||
| Phylum: | حبليات | ||
| الفرع الحيوي: | الأركوصورات التاجية | ||
| الفرع الحيوي: | الأركوصورات | ||
| الفرع الحيوي: | الطيور مشطية الأقدام | ||
| الفرع الحيوي: | طيريات الرقبة | ||
| الفرع الحيوي: | أشكال الديناصورات | ||
| الفرع الحيوي: | أشباه الديناصورات | ||
| الفرع الحيوي: | أشكال الديناصورات | ||
| الفرع الحيوي: | الديناصورات Owen, 1842 | ||
| Major groups | |||
| |||







الديناصورات (إنگليزية: Dinosaur، من الكلمة الإغريقية δεινόσαυρος، داينوسوروس)، هي حيوانات فقارية سادت النظام البيئي الأرضي لأكثر من 160 مليون سنة، بدءاً من العصر الثلاثي المتأخرة - قبل 230 مليون سنة - حتى نهاية العصر الطباشيري (قبل 65 مليون سنة) عندما انقرضت معظم الديناصورات في حدث الانقراض الطباشيري- الثلاثي. تُعتبر أنواع الطيور الحية اليوم من الديناصورات، بوصفها منحدرة من الديناصورات الثيروبودية.
تعتبر الديناصورات إحدى أكثر مجموعات مملكة الحيوان أشكالاً؛ فسليلتها من الطيور أكثر الفقاريات تنوعاً على وجه البسيطة، حيث يوجد منها ما يفوق 10,700 نوع.[7] استطاع علماء الأحياء القديمة التعرف على ما يزيد عن 500 جنس[8] و1,000 نوع مختلف من الديناصورات غير الطيرية،[9] واليوم يمكن العثور على أشكال مختلفة من الديناصورات في جميع قارات العالم، سواء حية ممثلة بالطيور، أو نافقة تُعرّف بواسطة مستحثاتها.[10] كان عدد من الديناصورات عاشباً وكان بعضها الآخر لاحماً، كما أن أنواعاً منها سارت على قائمتيها الخلفيتين، فيما سارت أنواع أخرى على أربع، واستطاع بعضها أن يتنقل باستخدام الطريقتين. طوّرت العديد من الديناصورات غير الطيرية امتدادات خارجية لهيكلها العظمي شكلت لها درعًا جسديًا أو طوقًا عظميًا أو قرونًا، وقد حققت بعض الفصائل شهرة عالمية بسبب غرابة شكلها الخارجي كما يتصوره العلماء. سيطرت الديناصورات الطيرية على أجواء الكرة الأرضية منذ انقراض الزواحف المجنحة التي كانت تعتبر منافستها الأساسية والسبب الرئيسي الذي يكبح تطورها. تشتهر الديناصورات بفعل حجم بعض الأنواع الضخمة، الأمر الذي يولد فكرة لدى العامّة مفادها أن جميع هذه الحيوانات كانت عملاقة، إلا أنه في واقع الأمر فإن معظم الديناصورات كانت بحجم الإنسان أو أصغر حتى. يُعرف أن معظم فصائل الديناصورات كانت تبني أعشاشًا لتضع فيها بيضها وتحضنه حتى الفقس. Their biology does not precisely correspond to the antiquated class Reptilia of Linnaean taxonomy, consisting of cold-blooded amniotes without fur or feathers. As Linnean taxonomy was formulated for modern animals prior to the study of evolution and paleontology, it fails to account for extinct animals with intermediate traits between traditional classes.}} of the clade Dinosauria. They first appeared during the Triassic period, between 243 and 233.23 million years ago, although the exact origin and timing of the evolution of dinosaurs is the subject of active research. They became the dominant terrestrial vertebrates after the Triassic–Jurassic extinction event 201.3 million years ago; their dominance continued throughout the Jurassic and Cretaceous periods. The fossil record shows that birds are modern feathered dinosaurs, having evolved from earlier theropods during the Late Jurassic epoch, and are the only dinosaur lineage to survive the Cretaceous–Paleogene extinction event approximately 66 million years ago. Dinosaurs can therefore be divided into avian dinosaurs, or birds; and the extinct non-avian dinosaurs, which are all dinosaurs other than birds.
Dinosaurs are a varied group of animals from taxonomic, morphological and ecological standpoints. Birds, at over 10,700 living species, are among the most diverse group of vertebrates. Using fossil evidence, paleontologists have identified over 900 distinct genera and more than 1,000 different species of non-avian dinosaurs. Dinosaurs are represented on every continent by both extant species (birds) and fossil remains. Through the first half of the 20th century, before birds were recognized as dinosaurs, most of the scientific community believed dinosaurs to have been sluggish and cold-blooded. Most research conducted since the 1970s, however, has indicated that dinosaurs were active animals with elevated metabolisms and numerous adaptations for social interaction. Some were herbivorous, others carnivorous. Evidence suggests that all dinosaurs were egg-laying; and that nest-building was a trait shared by many dinosaurs, both avian and non-avian.
While dinosaurs were ancestrally bipedal, many extinct groups included quadrupedal species, and some were able to shift between these stances. Elaborate display structures such as horns or crests are common to all dinosaur groups, and some extinct groups developed skeletal modifications such as bony armor and spines. While the dinosaurs' modern-day surviving avian lineage (birds) are generally small due to the constraints of flight, many prehistoric dinosaurs (non-avian and avian) were large-bodied—the largest sauropod dinosaurs are estimated to have reached lengths of 39.7 متر (130 أقدام) and heights of 18 m (59 ft) and were the largest land animals of all time. The misconception that non-avian dinosaurs were uniformly gigantic is based in part on preservation bias, as large, sturdy bones are more likely to last until they are fossilized. Many dinosaurs were quite small, some measuring about 50 سنتيمتر (20 بوصات) in length.
The first dinosaur fossils were recognized in the early 19th century, with the name "dinosaur" (meaning "terrible lizard") having been coined by Sir Richard Owen in 1841 to refer to these "great fossil lizards". Since then, mounted fossil dinosaur skeletons have been major attractions at museums worldwide, and dinosaurs have become an enduring part of popular culture. The large sizes of some dinosaurs, as well as their seemingly monstrous and fantastic nature, have ensured their regular appearance in best-selling books and films, such as Jurassic Park. Persistent public enthusiasm for the animals has resulted in significant funding for dinosaur science, and new discoveries are regularly covered by the media.
أصبحت الهياكل العظمية للديناصورات المعروضة في المتاحف حول العالم تشكل معالم جذب سياحية مهمة، منذ أن اكتُشف أول أحفور لديناصور في أوائل القرن التاسع عشر، وتحوّلت هذه الحيوانات إلى رمز من رموز الثقافة العالمية والمحلية لبعض البلدان. صُوّرت الديناصورات في الكثير من الروايات والأفلام السينمائية التي حققت نحاجًا كبيرًا ونسبة مبيعات هائلة، مثل سلسلة [[حديقة الديناصورات (فيلم)|حديقة الديناصوراتي]، كذلك فإن أي اكتشاف جديد على درجة كبيرة من الأهمية تغطيه وسائل الإعلام المختلفة وتقدمه للجمهور المهتم بهذه الأمور.
التسمية

صاغ سير ريتشارد اوين مصطلح "ديناصور" في 1842 من الجذور الإغريقية δεινός (داينوس) بمعنى: رهيب أو قوي أو مذهل، وσαῦρος (سوروس) بمعنى "عظاءة". يُستخدم المصطلح بشكلٍ غير علمي أحياناً لوصف زواحف قبل تاريخية أخرى مثل البليكوصور ديميترودون، والتيروصور المجنح، والإكثيوصور المائي، والبليسيوصور، والموساصور، برغم أن أياً من هذه الحيوانات لا ينتمي إلى طبقة الديناصورات. خلال النصف الأول من القرن العشرين اعتقد معظم المجتمع العلمي أن الديناصورات كانت حيواناتٍ بطيئة وغير ذكية وباردة الدم. مع ذلك، دعمت البحوث التي أجريت مُنذ السبعينيات المعتقد الذي يرى أن الديناصورات كانت حيواناتٍ نشيطة وتمتعت بأيضٍ مرتفع وتأقلمت بأشكالٍ مختلفة لتتواصل فيما بينها. وبشكلٍ تدريجي انتقل الفهم العلمي الجديد للديناصورات إلى الوعي الشعبي.
اقترح اكتشاف الطائر البدائي أركيوپتركس في 1861 وجود علاقة قريبة بين الديناصورات والطيور. وبغض النظر عن وجود طبعات ريش أحفورية، فإن الأركيوبتريكس كان شديد الشبه بمعاصره الديناصور المفترس كومپسوگناثوس. منذ ذلك الحين، رجحت البحوث كون الديناصورات الثيروپودية أسلاف الطيور المعاصرة، ويعتبر مُعظم علماء الإحاثة اليوم الطيور الديناصورات الوحيدة الناجية من الانقراض، ويقترح البعض وجوب تجميع الديناصورات والطيور في تصنيف بيولوجي واحد.[11] علاوة على الطيور، فإن القريب الآخر الوحيد الناجي إلى الزمن الحاضر للديناصورات هو التمساحيات. ومثل الديناصورات والطيور، فإن التمساحيات أعضاء في مجموعة أركوصوريا التي تضم زواحف العصر البرمي المتأخر التي حكمت الأرض الوسطى.
منذ اكتشاف أول مستحاثة ديناصور في أوائل القرن التاسع عشر، أصبحت هياكل الديناصورات العظمية نقاطٍ جذب هائلة للمتاحف حول العالم. صارت الديناصورات جزءاً من ثقافة العالم وحافظت على شعبيتها. ظهرت الديناصورات في أفلامٍ وكتبٍ حققت أعلى المبيعات (خصوصاً الحديقة الجوراسية)، وتغطي وسائل الإعلام الاكتشافات الحديثة في مجال الديناصورات بشكلٍ مستمر.
التعريف


تعريف عام

الديناصور حيوان زاحف عاش قبل ملايين السنين. وكلمة الديناصور مشتقة من كلمتين يونانيتين تعنيان السحلية المزعجة الرهيبة. ولم تكن الديناصورات سحالي ولكن حجم بعضها كان مروِّعًا، حيث إن الكبيرة منها كانت أضخم الحيوانات التي سكنت على اليابسة على الإطلاق، وكان وزنها أكثر من عشرات أضعاف وزن فيل كامل النمو. وهناك أنواع نادرة من الحيتان تنمو لتصبح أضخم من هذه الديناصورات.

ظهر أول الديناصورات على الأرض قبل مايقارب 220 مليون سنة مضت. وسيطرت هذه المخلوقات على اليابسة لما يقارب 150 مليون سنة، حيث عاشت في معظم بقاع العالم وفي أوساط متنوعة، من المستنقعات إلى السهول المنبسطة، إلا أنها انقرضت فجأة قبل نحو 63 مليون سنة.
وقد اختلفت الديناصورات بدرجة كبيرة في حجمها ومظهرها وعاداتها إلا أن أشهر أنواعها تشمل عمالقة مثل الأباتوسورس، والدبلودوكس، والتيرانوسورس. وإلاباتوسورس الذي سُمِّيَ أيضا بالبرونتوسورس ويصل طوله نحو 21م، في حين أن الدبلودوكس قد ينمو إلى أطول من ذلك، حيث بلغ نحو 27م، وقد كان كلاهما من آكلات النباتات. ولهذه الوحوش رؤوس صغيرة ورقاب وذيول طويلة للغاية. وكان التيرانوسورس من المفترسات آكلات اللحوم. وهو يقف بارتفاع ثلاثة أمتار عند الورك وله رأس ضخم وأسنان طويلة ومستدقة الأطراف. ولكن الديناصورات لم تكن جميعها عملاقة حيث كان أصغر أنواعها بحجم الدجاجة. وبصورة عامة فإن الديناصورات كانت تشبه معظم زواحف يومنا الحاضر. فعلى سبيل المثال، كان لبعضها أسنان وعظام وجلد مثل ما لدى التماسيح وبعض الزواحف الأخرى المعاصرة. ومن المحتمل جدًا أن بعضها كان يتمتع بدرجة ذكاء كتلك التي عند التماسيح، إلا أن الديناصورات اختلفت عن زواحف يومنا الحاضر بأساليب أخرى. فعلى سبيل المثال، لم ينَْمُ أي من الزواحف الحديثة بحجم كبير يشبه كبر الديناصورات. وهناك اختلاف آخر مهم وهو شكل وقفة الديناصور. ففي حالة السحالي والسلاحف ومعظم الزواحف الأخرى، فإن السيقان تكون ظاهرة على جانبي الجسم، حيث إن تركيب السيقان يُعطي هذه الحيوانات شكلاً منبسطًا وغير متناسق، في حين أن سيقان الديناصور تبقى تحت الجسم مشابهة بذلك سيقان الحصان. وقد أدى تركيب الساق هذا إلى رفع جسم الديناصور عن الأرض ومكن بعض أنواعها من السير على سيقانها الخلفية.
عاشت الديناصورات في فترة زمنية من تاريخ الأرض نسميها حقب الحياة المتوسطة أو الدهر الوسيط والتي استمرت من نحو 240 إلى 63 مليون سنة. وتسمى حقب الحياة المتوسطة أيضًا بعصر الزواحف حيث سيطرت الزواحف على الأرض والبحر والسماء أثناء تلك الفترة الزمنية. وانتمت أكثر الزواحف أهمية لمجموعة من الحيوانات سميت الأركوسورس (الزواحف السائدة). وبالإضافة إلى الديناصورات، شملت هذه المجموعة الثيكودونتس وهي أسلاف الديناصورات والتماسيح والزواحف الطائرة. وبنهاية حقب الحياة المتوسطة، فإن جميع الأركوسورس قد انقرضت فيما عدا التماسيح، وبذلك يكون عصر الزواحف قد انتهى.
ولم يعرف العلماء سبب اختفاء الديناصورات ولكنهم اعتقدوا أنها لم تترك سلالة. غير أن بعض علماء اليوم يعتقدون أن ديناصورات صغيرة معينة من آكلة اللحوم كانت هي أجداد الطيور.
عرف العلماء الديناصورات من خلال دراسة أحافيرها وهي عظامها وأسنانها وبيضها وآثار أقدامها المحفوظة، وهم يقومون أيضًا بدراسة الزواحف الحية والحيوانات الأخرى التي تمتلك صفات مشابهة لتلك الديناصورات.
تعريف معاصر
وفقاً لتصنيف النشوء فإن الديناصورات تُعرف عادة بأنها المجموعة التي تحتوي على "تريسراتبس، نيورنيثيس [الطيور المعاصرة]، أقرب أسلافها، وكل المتحدرين منها".[12] كما اقترح أن الديناصورات ينبغي أن تُعرف من وجهة كونها أقرب سلفٍ شائع للميگالوسوروس والإگواندون لأن هذين نوعان من ثلاثة نص عليها ريتشارد أوين عندما ميز الديناصورات.[13] ينتج عن التعريفين نفس مجموعة الحيوانات المعرفة بكونها ديناصورات، ومن ضمنها الثيروبودات (غالباً لواحم ثنائية القدم)، الصوروبودومورفات (غالباً عواشب رباعية القدم)، الأنكيلوصوريات (عواشب مدرعة رباعية القدم)، الستگوصوريات (عواشب مسطحة رباعية القدم)، السيراتوبسيات (عواشب رباعية القدم بقرون وأهداب، والأورنيثوبودات (عواشب ثنائية أو رباعية القدم من ضمنها "بطية المنقار"). كُتبت هذه التعريفات لتستجيب للمفاهيم العلمية حول الديناصورات التي تسبق الاستخدام الحديث للتطور العرقي. ويُقصد باستمرارية المعنى منع أي التباس بشأن معنى المصطلح "ديناصور".
يوجد شبه إجماع عالمي بين علماء الإحاثة على كون الطيور سليلة الديناصورات الثيرابودية. وباستخدام تعريف علم الأفرع المحدد الذي يوجب تضمين كل سلالات سلفٍ واحد شائع في مجموعة واحدة لجعل هذه المجموعة طبيعية، فإن الطيور ديناصورات، وبناء على ذلك فإن الديناصورات غير منقرضة. تصنف الطيور بواسطة معظم علماء الإحاثة على أنها تنتمي إلى المجموعة الفرعية مانيرابتورا، التي تنتمي إلى الكويلوروصوريا، التي تنتمي إلى الثيرابودات، التي تنتمي إلى الصوريسكيات، التي هي عبارة عن ديناصورات.[14]
من وجهة نظر علم الأفرع، الطيور ديناصورات، لكن كلمة "ديناصور" في الخطاب الاعتيادي لا تتضمن الطيور. بالإضافة إلى ذلك، فإن الإشارة إلى الديناصورات التي ليست طيوراً بوصفها "ديناصورات غير طيرية" مرهقة. ولأغراض التوضيح، ستستخدم هذه المقالة كلمة "ديناصور" كمرادف لمصطلح "الديناصورات غير الطيرية" الذي سيستخدم متى ما اقتضت الحاجة التوكيد. كذلك، فإنه من الصحيح تقنياً الإشارة إلى الديناصورات بوصفها مجموعة مميزة تحت نظام تصنيف لينيوس الأقدم، الذي يقبل التصنيف شبه العرقي الذي يستثني بعض السلالات المتحدرة من سلفٍ واحد شائع.
وبالرغم من أنها تعرف بحجمها الهائل، فإن كثيراً من الديناصورات كانت بحجم الإنسان أو أصغر. وُجدت بقايا الديناصورات في كُل القارات الأرضية، بما فيها القارة القطبية.[10] لا تُعرف ديناصورات عاشت في بيئة مائية أو هوائية برغم أن الثيرابودات المريشة يُمكن أن تكون طائرة.
الخصائص التشريحية المميزة


باستخدام التعريفات أعلاه، فإن الديناصورات (وليس من ضمنها الطيور) يُمكن أن توصف بأنها زواحف أركوصورية أرضية بأطرافٍ تنتصب تحت الجسم وُجدت مُنذ العصر الثلاثي المتأخر (حيث ظهر في الطبعات الصخرية) حتى العصر الطباشيري المتأخر (الذي انقرض في نهايته).[15] تُعتبر كثيرٌ من الحيوانات قبل التاريخية ديناصوراتٍ في التصور الشعبية، ومثالٌ على ذلك الإكثيوصورات، الموساصورات، البليسيوصورات، البتيروصورات، والديميترودون، لكن هذه الحيوانات لا تُصنف علمياً على أنها ديناصورات. الزواحف المائية كالإكثيوصورات والموساصورات والبليسيوصورات لم تكن مخلوقاتٍ أرضية أو أركوصورات، وكانت البتيروصورات أركوصورات لكنها لم تكن أرضية، بينما كان الديميترودون حيواناً برمياً أقرب إلى الثدييات. [16] كانت الديناصورات الفقاريات المهيمنة على النظام الأرضي في الحقبة الوسطى، خصوصاً خلال العصر الجوراسي والطباشيري. كانت مجموعات الحيوانات الأخرى خلال هذه الحقبة مقيدة من حيث الحجم والبيئة الملائمة، وعلى سبيل المثال، فنادراً ما تجاوزت الثدييات حجم قطة، وكانت غالباً لواحم بحجم القوارض تتغذى على فرائس صغيرة.[17] يُستثنى من هذه الحالة الربنومام العملاق: ترايكونودونت يزن ما بين 12 إلى 14 كيلوغراماً عُرف بأكله الديناصورات الصغيرة مثل السيتكوصورات الصغيرة.[18]
كانت الديناصورات مجموعة حيوانية شديدة التنوع، ووفقاً لدراسة أجريت في 2006 فإن خمسمائة (500) نوعٍ من الديناصورات قد عُرِفَ بدرجة كبيرة من التأكد حتى الآن، ويُقدر عدد الأنواع المحفوظات في السجلات الأحفورية بحوالي ألف وثمانمائة وخمسين (1850) نوعاً، يبقى حوالي 75% منها بانتظار أن تُكتشف. [19] وتنبأت دراسة سابقة بوجود ثلاثة آلافٍ وأربعمائة (3400) نوعٍ من الديناصورات، بينها مجموعة كبيرة لم تُحفظ في سجلات أحفورية.[20] واعتباراً من 17 سبتمبر 2008 سُمي ألف وأربعة وسبعون (1074) نوعاً من الديناصورات.[21] بعض هذه الديناصورات عاشب وبعضها الآخر لاحم، كما أن بعضها ثنائي القدم، وبعضها رباعي القدم، وبعضٌ ثالث كالأموصور والإغواندون كان يمشي على قدمين وعلى أربع أقدامٍ بسهولة. كان لبعضها درع عظمي، أو تعديلات جمجمية كالقرون.
بالرغم من أن الاكتشافات الحديثة صعبت تقديم قائمة متفق عليها بخصائص الديناصورات المظهرية المميزة، إلا أن كل الديناصورات المكتشفة حتى الآن تتشاطر تحوراتٍ معينة طرأت على هيكل الأركوصورات العظمي الأصلي. ومع أن بعض مجموعات الديناصورات اللاحقة احتوت تحوراتٍ إضافية، فإن التحورات الأساسية تعتبر نمطية في الديناصورات، حيث احتوت عليها الديناصورات المبكرة ونقلتها إلى كل المتحدرين منها.
صفات الديناصورات المشتركة تتضمن عرفاً مستطيلاً على العضد أو عظم الذراع العلوي لتحمل عضلات الكتف والساعد، بالإضافة إلى رف في مؤخرة عظم الورك، وحافة عريضة منخفضة قرب المؤخرة، وعظام كاحل مدعمة لتحمل الأطراف السفلية.[22]
التاريخ التطوري
الأصول والأسلاف


اعتقد العلماء لفترة طويلة أن الديناصورات متعددة الأعراق، حيث كان منها العديد من المجموعات غير المرتبطة، التي تشابهت مع غيرها في بعض الأحيان كونها تعرّضت لظروف بيئية مشابهة دفعتها لتتطور في نمط متشابه.[23][24][25] إلا أن الاكتشافات الحديثة أظهرت أن كل أنواع وفصائل الديناصورات تُشكل معًا مجموعة واحدة غير متفرعة.[12][26]
انشقت الديناصورات عن أسلافها من الأركوصورات منذ حوالي 230 مليون سنة، أي منذ أواسط حتى أواخر العصر الثلاثي، وبهذا فإنها تكون قد ظهرت بعد 20 مليون سنة من حادثة انقراض العصر البرمي-الثلاثي، التي قضت على ما يقرب من 95% من أشكال الحياة على الأرض،[27][28] وقد دعمت الدراسات الإشعاعية لتحديد عمر طبقات الصخور التي حوت المستحثات الخاصة بجنس الديناصور الفجري أيورابتور، وهو أحد أكثر أجناس الديناصورات بدائيةً، دعمت هذه النظرية سالفة الذكر.[29] يقول علماء الأحياء القديمة أن الأيورابتور يشابه السلف المشترك لكل الديناصورات؛ وإن ثبتت صحة هذه النظرية فإن هذا يفيد بأن الديناصورات البدائية كانت مفترسات صغيرة الحجم ذات قائمتين.[30] أظهرت بعض الاكتشافات لديناصورات طيرية العنق في طبقات العصر الثلاثي بالأرجنتين أن هذه الحيوانات كانت بالفعل مفترسات صغيرة ذات قائمتين، الأمر الذي يدعم النظرية سالفة الذكر.
كانت الموائل الأرضية مأهولة بأنواع عديدة من الأركوصورات الأولى والزواحف شبيهة الثدييات خلال الزمن الذي ظهرت فيه الديناصورات، ومن هذه الكائنات: السحالي العقابية أو الأيتوصورات (باللاتينية: Aetosauria)، وكلبيات الأسنان (باللاتينية: Cynodontia). انقرضت معظم هذه الحيوانات أثناء العصر الثلاثي في حدثيّ انقراض كبيرين، فمن المعروف أنه خلال القسمين الثاني والثالث من الدور الأول للعصر الثلاثي، أي منذ حوالي 215 مليون سنة، انقرض الكثير من تلك الكائنات البدائية، ومن ثم وقعت حادثة انقراض أخرى منذ حوالي 200 مليون سنة هي حادثة انقراض العصر الثلاثي-الجوراسي، وفيها اندثر ما تبقى من الأركوصورات البدائية، وخلت الأرض لشبيهات التماسيح والديناصورات والثدييات والزواحف المجنحة والسلحفيات.[12] أخذت الديناصورات تتنوع وتتفرع لفصائل مختلفة وأشكال وأحجام عديدة خلال المراحل الأخيرة من العصر الثلاثي، وذلك عن طريق سيطرتها على الموائل الطبيعية التي فرغت من قاطنتها، ولعبها لذات الدور البيئي التي لعبته الحيوانات السابقة لها. كان العلماء يعتقدون أن الديناصورات ساهمت في تسريع وتيرة انقراض الحيوانات البرية الأخرى التي عاصرتها خلال العصر الثلاثي بسبب أن الأخيرة لم تقو على منافستها، فتناقصت أعدادها كنتيجة لهذا، وحلت الديناصورات بدلاً منها، إلا أن هذه النظرية تبدو الآن أقل واقعيةً لأسباب مختلفة: فسجل المستحاثات لا يُظهر نموًا سريعًا وازدهارًا كبيرًا في أعداد وفصائل الديناصورات خلال تلك الفترة كما توقع العلماء؛ بل يظهر بأنها كانت لا تزال نادرة في بداية الدور الأول من أواخر العصر الثلاثي حيث تراوحت نسبتها بين 1% و 2% من إجمالي الحيوانات التي كانت موجودة آنذاك، أما في الدور الثاني من أواخر ذلك العصر، تفيد المستحاثات أن أعدادها قد ارتفعت وفصائلها قد إزدادت، حيث ترواحت نسبتها بين 50% و 90% من إجمالي الحيوانات. كذلك كان العلماء يقولون أن الوضعية المنتصبة للديناصورات البدائية كانت عاملاً مساعدًا جعلها تتفوق على معظم الكائنات الأخرى رباعية القوائم، إلا أنه يُعرف الآن أن هذه الخاصية كانت موجودة عند الكثير من الحيوانات التي عاصرت أوائل عهد الديناصورات، ولم تكن سمة مميزة لمجموعة كائنات واحدة. أخيرًا فإن الأحافير تُظهر انفجارًا في أشكال الحياة حصل في أواخر العصر الثلاثي وأدى إلى بروز بعض مجموعات الحيوانات،[12] مثل مجموعة "مصلبة الكاحلين" (باللاتينية: Crurotarsi) التي لم يبق من رتبها وصفوفها اليوم إلا التمساحيات، التي كانت أكثر تنوعًا من الديناصورات بكثير، وبحسب الظاهر فإن هكذا مجموعات متنوعة وكبيرة من شأنها أن تهيمن على المجموعات الأقل عددًا وتنوعًا، لذا فإن نشوء وارتقاء الديناصورات حتى أصبحت هي الكائنات المسيطرة على الأرض كان بفعل "الحظ" أكثر من تفوقها، كما يفيد العلماء من أنصار هذا الاتجاه.[31]
تراجع التنوّع خلال العصر الطباشيري
تفيد التحليلات الإحصائية المستندة إلى بعض المعلومات غير المدروسة، إلى أن الديناصورات ازداد تنوعها خلال أواخر العصر الطباشيري، حيث ظهرت فيها أجناس جديدة تفرعت إلى فصائل وأنواع كثيرة. إلا أنه في عام 2008، قال بعض العلماء أن هذا الكلام غير دقيق، وإن ما قيل حول تنوع الديناصورات في تلك الفترة إنما هو نتيجة خطأ علمي، وأفادوا أن الديناصورات لم يظهر فيها سوى مجموعتين جديدتين في أواخر العصر الطباشيري هما بطيّات المنقار أو الهادروصورات، وذوات القرون أو السحالي القرناء. يفيد العلماء أن ما كبح ظهور مجموعات جديدة من الديناصورات هو تطوّر النباتات، ففي أواسط العصر الطباشيري أصبحت النباتات المزهرة مغطاة البذور جزءًا مهمًا من النظام البيئي العالمي، وانتشرت على سطح الكرة الأرضية مستبدلة عاريات البذور مثل الصنوبريات، ويدل براز الديناصورات المتحجر أن هذه الكائنات استمرت تقتات على النباتات عارية البذور بشكل أكبر من مغطاة البذور، فيما تحوّلت الكثير من الحشرات النباتية والثدييات إلى الاقتيات على مصدر الغذاء الجديد، وفي الوقت نفسه حصل انفجار في أنواع السحالي والأفاعي والتمساحيات والطيور، فأخذت فصائل جديدة منها تظهر وتزدهر وتحل مكان الديناصورات شيءًا فشيئًا. يقول بعض العلماء أن فشل الديناصورات في التنوع والارتقاء في الوقت الذي كان فيه النظام البيئي حولها يتغيّر، حكم عليها بالانقراض بعد بضعة ملايين السنين.[32]
التطور
ارتبط تطور الديناصورات بعد نهاية العصر الثلاثي بتطور النباتات وبزحل القارّات، ففي أواخر ذاك العصر وعند بداية العصر الجوراسي، كانت جميع قارّات العالم متصلة ببعضها البعض وتُشكل قارة عظمى تُعرف باسم قارة "بانجيا"، وكانت جمهرة الديناصورات مكونة بمعظمها من اللواحم الحلقية وطويلات العنق الأولية العاشبة،[33] كذلك سيطرت على الأرض غابات من الأشجار عارية البذور، وبشكل خاص الصنوبريات، التي شكلت غذاءً بارزًا للعواشب. افتقدت طويلات العنق الأولية إلى الآليات المعقدة اللازمة في الفم لمعالجة أوراق الصنوبريات الإبريّة، لذا كانت تبتلعه كاملاً، ويُحتمل أن تكون قد امتلكت بعض الخاصيّات الأخرى في قناتها الهضميّة لتجزئة الطعام وتكسيره إلى العناصر الغذائية اللازمة لبقائها.[34] استمرت الديناصورات متجانسة ومتماثلة طيلة أواسط وحتى أواخر العصر الجوراسي، حيث كانت المفترسات من السيراتوصوريات والسبينوصوريات والكارنوصوريات تنتشر في مختلف المناطق، إلى جانب عواشب ضخمة ممثلة بالديناصورات المغطاة أو الستيغوصوريات وطويلات العنق أو الصوروبوديات. ومن الأماكن التي تُظهر هذا التجانس في أنواع وأجناس ديناصوراتها على الرغم من بعدها عن بعضها حاليًا: تكوين موريسون في ولاية كولورادو في الولايات المتحدة، وتكوين تنداجورو في تنزانيا. وحدها ديناصورات الصين من تلك الحقبة تُظهر اختلافًا عن باقي الديناصورات التي عاصرتها، حيث كان منها لواحم ثيروبوديّة متخصصة في أنواع معينة فقط من الطرائد، وطويلات عنق لا نمطيّة مثل ديناصور مامنتشي مامنتشيصور.[33] أخذت الديناصورات المنصهرة، أو الأنكيلوصوريات، تصبح أكثر شيوعًا خلال أواخر العصر الجوارسي، وكذلك الحال بالنسبة للديناصورات طيرية الورك، وبحلول ذلك الوقت كانت طويلات العنق الأوليّة قد انقرضت، وأصبحت الصنوبريات والسراخس أكثر النباتات انتشارًا. كانت طويلات العنق غير قادرة على معالجة الطعام في فمها كما كان حال أسلافها، إلا أن الديناصورات طيرية الورك كانت قد طوّرت مقدرةً على تقطيع الطعام ومضغه في فمها قبل أن تبتلعه، وذلك عن طريق ظهور أفكاك متحركة لها،[34] لكن لعلّ أبرز حدث تطوريّ وقع خلال العصر الجوراسي هو ظهور الطيور الحقيقية، التي تطورت من اللواحم الحلقية.[14]

أخذت الديناصورات تتفرع لأشكال أكثر تنوعًا خلال أوائل العصر الطباشيري بسبب تفكك قارة بانجيا وانعزال المزيد من الجمهرات عن مثيلتها. كانت الديناصورات المنصهرة إلى جانب ذوات الأسنان الإغوانية والديناصورات الذراعية أو البراكيوصوريات، هي أول المجموعات التي انتشرت خلال فجر هذا العهد وتوزعت في مختلف أنحاء أوروبا وأمريكا الشمالية وشمال أفريقيا، وفي وقت لاحق استبلدت اللواحم الجوارسية الأفريقية بلواحم أوائل العصر الطباشيري العملاقة، مثل السبينوصور والكاركارادونتوصور، وورثت أجناس جديدة من طويلات العنق مواطن الصوروبودات الجوارسية، ومن هذه الأجناس: الريباكيصور والتياتانوصور، التي عاشت في أمريكا الجنوبية إلى جانب أفريقيا. ظهرت في آسيا خلال هذه الفترة فصائل وفيالق جديدة من اللواحم الحلقية، لعلّ أبرزها هو الديناصورات السريعة أو الدروميوصوريات، المعروفة أكثر باسم "الكواسر"، كذلك ظهرت الترودونيات وكواسر البيض أو الأوڤيرابتورات، وأصبحت هذه مجتمعةً تُشكل مجموعة اللواحم الأساسية. أيضًا انتشرت الأنكيلوصوريات وبرزت ذوات قرون بدائية مثل الديناصور الببغائي سيتكوصور، وأصبحت إحدى أبرز العواشب.[33] تُظهر المستحاثات أن الديناصورات المغطاة اختفت من على وجه الأرض خلال أواخر الدور الأول من العصر الطباشيري أو خلال أوائل الدور الأخير منه، كذلك يتبين أن النباتات المزهرة كانت قد نشأت وبدأت بالانتشار على نطاق شديد الضيق. طوّرت فصائل عديدة من الديناصورات وسائل مختلفة لمعالجة الطعام قبل ابتلاعه خلال أوائل العصر الطباشيري، فذوات القرون مثلاً طوّرت المقدرة على تقطيع النباتات بأسنانها المكدسة في صفوف، وطوّرت ذوات الأسنان الإغوانية المقدرة على طحن النبات بأسنانها، وتفوّقت في هذا المجال بطيّة المنقار.[34] ظهرت لدى بعض طويلات العنق صفوف من الأسنان خلال هذا العصر أيضًا، ومن أبرز الأمثلة على ذلك ديناصور النيجير نيجيرصور.[35]


سيطرت 3 فصائل كبرى من الديناصورات على مناطق مختلفة من العالم خلال أواخر العصر الطباشيري، ففي قارة "لوراسيا"، التي تكونت من أمريكا الشمالية وآسيا، كانت الديناصورات الطغاة أو التيرانوصوريات بالإضافة لعدّة أنواع من الكواسر ذات الأيادي السالبة هي الضواري الثيروبوديّة السائدة، وعاشت إلى جانبها مجموعة كبيرة من العواشب طيرية الحوض تمثلت ببطيّة المنقار وذوات القرون والديناصورات المنصهرة وسميكة الرأس. أما في القارة الجنوبية أو قارة "غندوانا" فكانت الديناصورات الأبيليّة أو الأبيليصورات هي الثيروبودات المهيمنة، وكانت التيتانوصوريات هي أكثر العواشب انتشارًا. كذلك عاشت الدروميوصوريات إلى جانب فصائل مختلفة من ذوات الأسنان الإغوانية والأنكيلوصوريات والتيتانوصوريات في أماكن مختلفة من أوروبا.[33] كانت النباتات المزهرة قد أصبحت واسعة الانتشار بحلول هذه الفترة من الزمن،[34] وأخذت الأعشاب البدائية بالظهور في نهاية العصر الطباشيري،[36] وكنتيجة لهذا التنوع في الغطاء النباتي، ازدهرت بطيّة المنقار وذوات القرون وظهرت منها أنواع مختلفة في جميع أنحاء أمريكا الشمالية وآسيا، وبطبيعة الحال ظهرت أنواع جديدة من اللواحم الثيروبوديّة لتغتنم هذا التنوع في الطرائد، وإلى جانبها ظهرت ديناصورات قارتة فريدة من نوعها، مثل الديناصورات البليدة والديناصورات النعاميّة المعروفة أيضًا باسم "الديناصورات محاكية الطيور" (باللاتينية: Ornithomimosauria).[34]
اندثرت الديناصورات من على وجه الأرض بعد وقوع حدث انقراض العصر الطباشيري-الثلاثي، منذ حوالي 65 مليون سنة في نهاية العصر الطباشيري، ولم ينج منها أي نوع أو أصنوفة سوى الطيور. كذلك نجت بعض الزواحف الأخرى ذات الجماجم القنطريّة، مثل التمساحيات، السحالي، الأفاعي، السحالي شبيهة الزواحف، وشبيهات الغريال.[37]
التصنيف
تُصنّف الديناصورات، بما فيها الطيور، على أنها أركوصورات ومثلها كمثل الزواحف. وتتميز الأركوصورات بجماجم "قنطريّة"، أي أنها تمتلك فجوات أو "نافذات" حرارية قنطرية أو قوسيّة في الموقع الذي تتصل فيه عضلات الوجه بالجمجمة، بالإضافة إلى نافذة أخرى قبمحجرية تقع في مواجهة العينين، وهذه السمات مشتركة بين الديناصورات ومعظم الزواحف المعاصرة والطيور. تتشارك الديناصورات في سمات تشريحية عديدة أخرى مع غيرها من الأركوصورات، لعلّ أبرزها هو الأسنان التي تنمو من أغمد عوض أن تكون امتداد مباشر لعظم الفك، وبالمقابل فهي تختلف عن غيرها من الأركوصورات بشكل بارز من ناحية مشيتها، فقوائم الديناصورات تمتد مستقيمةً أسفل جسدها، في حين أن قوائم السحالي والتمساحيات مفلطحة، تبدو للناظر وكأنها تنبثق من جانبها.
-
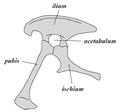 Saurischian pelvis structure (left side)
Saurischian pelvis structure (left side) -
 Tyrannosaurus pelvis (showing saurischian structure – left side)
Tyrannosaurus pelvis (showing saurischian structure – left side) -
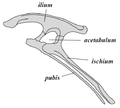 Ornithischian pelvis structure (left side)
Ornithischian pelvis structure (left side) -
 Edmontosaurus pelvis (showing ornithischian structure – left side)
Edmontosaurus pelvis (showing ornithischian structure – left side)




يُصنف عدد من العلماء الديناصورات كطبقة أو فرع حيوي آخر غير مُسمّى بعد. تُقسم الديناصورات إلى رتبتين: سحلية الحوض وطيرية الحوض، وفقًا لبنية حوضها. تضم رتبة سحلية الحوض تلك الأصنوفات التي تتشارك في أحد أسلافها مع الطيور المعاصرة عوض طيرية الحوض، وتضم الأخيرة جميع الأصنوفات التي تتشارك في أحد أسلافها مع الديناصور ثلاثي القرون ترايسيراتوبس عوض سحلية الحوض. حافظت الديناصورات سحلية الحوض على بنية ورك أسلافها، حيث بقي عظم العانة مواجهًا للأمام،[38] على الرغم من أن بعض الفصائل والأنواع المنتمية لهذه الرتبة طوّرت بعض السمات الخاصة بها في وقت لاحق، مثل الديناصورات الهيراريّة (باللاتينية: Herrerasauridae)[39] والديناصورات البليدة (باللاتينية: Therizinosauroidea)[40] والديناصورات السريعة (باللاتينية: Dromaeosauroidea)[41] والطيور،[14] حيث أصبحت عانتها أضيق أو أوسع بدرجات مختلفة. تضم رتبة سحليات الحوض الديناصورات الثيروبودية وشبيهات طويلة العنق (باللاتينية: Sauropodomorpha).

تميزت الديناصورات طيرية الحوض بحوض شديد الشبه بحوض الطيور، حيث كانت عظام عانتها ذيلية الشكل تتجه نحو الخلف كما الطيور، كذلك تُظهر المستحاثات أن عددًا كبيرًا من هذه الحيوانات كان يمتلك زائدةً أو "حدبة" عظميّة تتجه نحو الأمام. ضمّت هذه الفئة من الديناصورات أنواعًا عديدة من العواشب الضخمة.
استنادًا إلى المدونة الدولية لتسميات الحيوان، يمكن القول أن ما بين 630 إلى 650 نوعًا من الديناصورات كانت تعيش على وجه البسيطة منذ ملايين السنين، وما زال العلماء يقومون باكتشافات جديدة كل سنة تضيف أنواعًا وفصائل جديدة إلى القائمة الموجودة. يُلاحظ أن أكثر من نصف الديناصورات المكتشفة غير ممثل إلا بعينة واحدة وغالبًا ما تكون غير مكتملة، كذلك فإن أقل من 20% من الأنواع المصنفة غير معروفة إلا عن طريق 5 عينات أو أقل حتى. يحوي الجدول التالي المجموعات الأساسية من الديناصورات بالإضافة لبعض الأجناس والفصائل:
| سحليّة الحوض |
| ||||||
| الديناصورات الثيروبودية |
| ||||||
| الثيروبوديات الحديثة |
| ||||||
| الديناصورات صلبة الذيل |
| ||||||
| |||||||
| شبيهات طويلة العنق |
| ||||||
| طويلات العنق |
| ||||||
| طويلات العنق الحديثة |
| ||||||
| |||||||
| طيريّة الحوض |
| ||||||
| الديناصورات الوجنيّة | المدرّعات |
| |||||
| عريضة الورك |
| ||||||
| |||||||
| قرناء الورك | مهدبة الرأس |
| |||||
| |||||||
| طيرية الورك |
| ||||||
| |||||||




أنواع الديناصورات

قسم العلماء الديناصورات إلى مجموعتين رئيسيتين:
1-الصُّوريشيان.
2- الأورنيثيسشيان.
وتختلف هاتان المجموعاتان في تركيب الورك: فالأولى التي يعني اسمها السحلية الوركية تمتاز بوجود ورك مشابه لورك السحالي، في حين أن الثانية تعني الطير الوركي ويمتاز بوجود ورك مشابه لورك الطيور. وكل مجموعة من هاتين المجموعتين تتكون من عدة سلالات أساسية من الديناصورات.
وقد عاشت بعض أنواع الديناصورات طوال حقب الحياة المتوسطة. في حين أن أنواعًا أخرى عاشت فقط في فترة أو فترتين من الفترات الثلاث التي تتكون منها الحقبة كلها. وهذه الفترات الثلاث هي العصور الترياسي والجوراسي والكريتاسي. وقد امتد عصر الترياسي فيما بين 240 و205 مليون سنة ونصف في حين أن عصر الجوراسي امتد فيما بين 205و 138مليون سنة مضت. أما عصر الكريتاسي (وهو العصر الأطول في حقبة الميسوزوي) فقد استمر تقريبًا فيما بين 138 – 63 مليون سنة خلت.
الصوريشيان
احتوت على أضخم وأشرس الديناصورات وكان هناك ثلاثة أنواع أساسية من الصوريشيان وهي:
1- البروسوروبود
2- السوروبود
3ـ الثيروبود
واشتملت كل مجموعة من هذه المجموعات على العديد من ديناصورات السحالي الوركية.
البروسوروبود. كما هو الحال في البلاتوسورس، نمت إلى أن بلغ طولها ستة أمتار وكانت تتميز بعنق طويل ورأس صغير. وكانت تسير على رجليها الخلفيتين أحيانًا وعلى أرجلها الأربع أحيانًا أخرى. وكانت هذه أول الديناصورات آكلة النباتات. وقد ظهرت قبل نحو 220 مليون سنة وعلى مايبدو فقد انقرضت في بداية العصر الجوراسي.
السوروبود. كانت عمالقة عالم الديناصورات. وبلغ طولها نحو 21م وارتفاعها من الورك عند جلوسها حوالي ثلاثة أمتار ونصف المتر إلى أربعة أمتار ونصف المتر،. وكان وزن معظم أفرادها الكاملة النمو يتراوح بين تسعة أطنان وسبعة وعشرين طنًا متريًا. وكانت السوروبود تمشي على أربع أرجل ضخمة كما هي الحال في الفيل. والسوروبود المثالي له رقبة طويلة ورأس صغير وذيل طويل وصدر ضخم وعميق. وكان السوروبود هو آكل النبات الرئيسي في العصر الجوراسي وقد أصبحت بعض آكلة النبات الأخرى أكثر أهمية في العصر الكريتاسي. وقد وجد مختصو الأحافير بقايا السوروبود في كل القارات فيما عدا القارة القطبية الجنوبية. ويُعْد الأباتوسورس أو البرونتوسورس (البرونتصورص) من أفضل السوروبود المعروفة. وبالنسبة للعديد من الناس، فإن كلمة ديناصور تستحضر للذهن صورة الأباتوسورس، حيث كانت سيقانه الأمامية أقصر من سيقانه الخلفية وكان ظهره منحدرًا تجاه قاعدة العنق. أما الدبلودوكس وهو أطول الديناصورات المعروفة فقد بدا مشابهًا بدرجة كبيرة للأباتوسورس إلا أنه أنحف وأخف وزنًا. ووصل نموه إلى نحو 27م. وعاش كل من الأباتوسورس والدبلودوكس خلال العصر الجوراسي فيما يعرف الآن بأمريكا الشمالية. وقد عثر على بقايا الاباتوسورس في أوروبا.
البراكيوسورس. وهو نوع آخر من السوروبود، عُرف هذا الحيوان الضخم المخيف في شرق إفريقيا فقط. ومع ذلك، فإن البراكيوسورس قد عاش في العديد من مناطق العالم خلال العصر الجوراسي. ووصل ارتفاع البراكيوسورس إلى 12م أو أكثر. وبلغ وزنه مايصل إلى 77طنًا متريًا. وكانت سيقانه الأمامية أطول من الخلفية. وكانت هذه الحيوانات تقف مثل الزرافات حيث ينحدر الظهر إلى الأسفل باتجاه الذيل.
الثيروبود. كانت الوحيدة من الديناصورات آكلة اللحوم . وسارت هذه الحيوانات منتصبة القامة على رجليها الخلفيتين. وديناصورات الثيروبود المثالية امتازت بذيل عضلي طويل تحمله خلفها بشكل مستقيم لحفظ التوازن. وكانت أطرافها الأمامية نحيفة. وللثيروبود الكبيرة عنق قصير ورأس طويل ضخم. أما الثيروبود الصغيرة فكانت لها أعناق طويلة ورؤوس أصغر. ولجميع الثيروبود فكوك قوية وأسنان تشبه الشفرات. وقد عاشت طوال الحقبة الميسوزوية.
والتيرانوسورس المفترس من أشهر الثيروبود المعروفة ويعني اسمها هذا ملك السحالي الطاغية وهو الأكثر رعبًا من بين آكلة اللحوم في عصره. ويبلغ طول التيرانوسورس ثلاثة أمتار من الورك وينمو طوله إلى 12م تقريبًا. وبلغ طول رأسه حوالي المتر طولاً وبلغت أسنانه حوالي 15سم طولاً. وكان للحيوان أطراف أمامية قصيرة جدًا لدرجة أنها قد تكون عديمة الفائدة. ولكل طرف أمامي أصبعان. وقد ساد التيرانوسورس اليابسة إلى قرابة نهاية عصر الزواحف. وقد اكتُشفت بعض الأحافير القليلة للتيرانوسورس في أمريكا الشمالية. ووجد بعض الأحافير للثيروبود الكبيرة في آسيا وأمريكا الجنوبية.
قبل حوالي 150 مليون سنة ولفترة طويلة قبل ظهور التيرانوسورس، كانت الألوصورات هي الديناصورات الرئيسية الآكلة للحوم. وقدكانت تشبه التيرانوسورس ولكنها لم تكن بنفس الضخامة. كما أمتازت الألوصورات بوجود أطراف أمامية أطول وبثلاثة أصابع في كل من هذه الأطراف. وشملت الثيروبود الصغيرة الدينونيكس والأورنيثوميس. وكان الدينونيكس حوالي المتر طولاً وفي كل قدم يوجد مخلب منحنٍ ضخم ربما كان يستخدمه لتمزيق فريسته. أما الأورنيثوميس فقد كان على شكل نعامة فاقدة الريش وبنفس حجمها تقريبًا. واشتملت الثيروبود على أصغر أنواع الديناصورات المعروفة وهو الكومسوجناثس حيث كان هذا الحيوان بحجم الدجاجة تقريبًا.
الأورنيثيسشيان
كانت من آكلة النبات. وكانت أسنانها مزودة بزوائد عظمية منقارية الشكل وللعديد منها صفائح عظمية في جلدها. وفي العصر الطباشيري (الكريتاسي)، أصبحت الأورنيثيسشيان أهم الديناصورات آكلة النبات. وكان هناك أربعة أنواع أساسية منها وكان ترتيبها حسب ظهورها على اليابسة كما يلي:
1- الأورنيثوبودس.
2- الستيجوسورس (الأسطغور).
3- الأنكيلوسورس.
4- السيراتوبسيان. وقد اشتمل كل نوع على أعداد مختلفة
الأورنيثوبودس وهو يستطيع السير إما على أربع أرجل وإما على الرجلين الخلفيتين فقط. وأولى الأحافير الديناصورية المكتشفة كانت للإجوانودون وهو أحد أنواع الأورنيثوبودس. وبلغ طول الإجوانودون حوالي تسعة أمتار طولاً. وكانت له شوكة عظمية في إبهام كل طرف من أطرافه الأمامية وعاشت الأورنيثوبود طوال عصر الزواحف.
ووصلت الأورنيثوبودس قمة تطورها في ديناصور الدَّكبيل أو الهادروسورس. وكانت ديناصورات الدَّكبيل هذه منتشرة بشكل كبير خلال العصر الطباشيري. وقد عاشت في المناطق المعروفة الآن بآسيا وأستراليا وأمريكا الشمالية. وكان لكل واحد منها منقار منبسط في مقدمة الفم يشبه منقار البط وبه فكوك بها مئات الأسنان تمتد إلى مؤخرة الفم. كما كانت لها سيقان خلفية قوية وطويلة، وأطراف أمامية نحيلة ذات أصابع شبكية. ونمت ديناصورات الدَّكبيل هذه إلى ما يقترب من مترين وسبعة أعشار المتر ارتفاعًا من الورك وبامتداد ما يزيد على تسعة امتار.
وهناك بعض أنواع ديناصورات الدَّكبيل مثل الأناتوسورس ذات الجمجمة المنبسطة والمنخفضة. وأنواع أخرى مثل الكوريثوسورس التي لها عُرف عظميً ظاهر في أعلى رأسها. ولها قنوات هوائية ممتدة من أنوفها خلال هذا العُرف. ويعتقد بعض العلماء أن ديناصورات الدَّكبيل العُرفية هذه ربما كانت تصدر أصواتًا مرعبة باستعمال القنوات الهوائية الشبيهة بالبوق.
الستيجوسورس كانت من آكلات النباتات الضخمة الكبيرة التي تحمل صفائح عظمية على ظهورها القائمة، وقد عاشت قبل نحو 150 مليون سنة مضت. ومن أحسن ما عُرف من هذا النوع هو الستيجوسورسْ الذي عاش فيما يُعرف الآن بأمريكا الشمالية. وقد سارت هذه على أربع أرجل وبلغ طولها نحو ستة أمتار وارتفاعها حوالي مترين ونصف المتر من الورك ولها رأس صغير ورقبة قصيرة. وكانت سيقانها الخلفية أطول كثيرًا من الأمامية. ولهذا فقد كانت رؤوس هذه الحيوانات متدلية بالقرب من الأرض وبدت وكأنها منحنية للأمام.[42]
وكان للستيجوسُورس صف أو صفَّان من الصفائح العظمية الرأسية الممتدة على طول الظهر. وكان ذيلها مسلحًا بزوجين من الأشواك العظمية. وربما ساعدت هذه الأشواك والصفائح في حمايته من الأعداء. وربما ساعدت هذه الصفائح أيضًا في العمل كجزء من نظام تبريد للجسم. فمن الممكن أن يقوم الهواء المار حول وفوق الظهر بتبريد الدم المار في هذه الصفائح.
الأنكيلوسورس (الأنكيلوصورات) وهي معروفة بالديناصورات المدرعة. وهي حيوانات قصيرة وعريضة وتمشي على أربع أرجل وقد تراوح طول معظم الأنكيلوسورس بين أربعة أمتار ونصف المتر وستة أمتار ولها جمجمة يزيد طولها على نصف المتر. وقد غطت أجسام ورؤوس معظم الأنكيلوسورس صفائح عظمية ثقيلة. والعديد من هذه الصفائح ذو أشواك أو زوائد. وتنمو هذه الأشواك عادة على الأكتاف أو على خلفية الرأس. ولبعض أنواع الأنكيلوسورس كتلة كبيرة من العظام في نهاية الذيل ويمكن استعمالها كعصا ضد الأعداء. وقد عاشت الأنكيلوسورس في العديد من مناطق العالم إبان العصر الكريتاسي.
السيراتوبسيات كانت ديناصورات ذات قرون، وكانت تمشي على أربع أقدام وشبيهة إلى حد ما بحيوان وحيد القرن ويتراوح طولها بين متر وثمانية أعشار المتر، وسبعة أمتار وستة أعشار المتر ولها رأسٌ ضخمٌ. كما كان للسيراتوبسيان هدب عظمي على الجزء الخلفي من الرأس. وامتد هذا الهدب عبر العنق. وفي أحد أنواع السيراتوبسيان مثل الستايراكوسورس، كانت هذه الأهداب مزودة بأشواك. ولمعظم هذه الأنواع قرونٌ على وجهها. فالترايسيراتوب لها ثلاثة قرون وجهية، واحد قصير على الأنف وواحد فوق كل عين. وتنمو القرون التي فوق عيونها لتصل في طولها إلى متركامل. والمنونكلونيس له قرن واحد كبير على الأنف. وللبنتاسيراتوب خمسة قرون على وجهه. وأحد أنواع السيراتوبسيان وهو البروتوسيراتوب كان عديم القرون. وكانت السيراتوبسيان آخر المجموعات الرئيسية من الديناصورات. وقد عاشت فيما هو معروف الآن بآسيا وأمريكا الشمالية.
الخصائص الأحيائية
يشتق علماء الإحاثة والأحياء القديمة معلوماتهم عن الديناصورات من مجموعة كبيرة من السجلاّت الأحفورية وغير الأحفورية، وهذه تشمل: العظام المتحجرة، البراز المتحجر، آثار الأقدام المحفوظة، حصاة المعدة، طبعات الريش والجلد الصخريّة، بالإضافة لبعض الأعضاء الداخلية والأنسجة الحيوية المحنطة تحنيطًا طبيعًا.[43][44] تساهم الكثير من الدراسات الأخرى غير الأحفورية بالمساعدة على فهم هذه الكائنات وأسلوب حياتها، ومن هذه الدراسات: الفيزياء وبشكل خاص دراسات الميكانيكا الحيوية، الكيمياء، علم الأحياء، وعلوم الأرض الأخرى غير علم الأحياء القديمة. كان حجم الديناصورات وسلوكها من أكثر المواضيع إثارة لاهتمام العلماء، لذا فإنها تعرضت للدراسة المكثفة وظهرت فيها نظريات عديدة خلال عقود من الزمن.
المعيشة

اعتقد الناس ولسنوات عديدة أن الديناصورات مخلوقات رعناء، بطيئة الحركة، وعاشت بشكل يشبه زواحف اليوم، إلا أن البراهين الأحفورية بينت أن بعض أنواع الديناصورات خصوصًا الثيروبود الصغيرة ربما كانت أنشط كثيرًا من معظم زواحف اليوم، إضافة إلى أن معظم الديناصورات أقرب شبهًا بالطيور أكثر من شبهها بزواحف اليوم، وذلك فيما يتعلق بسيقانها وبنية أقدامها وهيئتها المنتصبة. ويعتقد بعض العلماء أن الديناصورات أقرب بأن تكون أسلافًا للطيور منها للزواحف اليوم. ويعتقدون أيضًا أن دراسة الطيور يمكن أن تساعدنا على معرفة حياة الديناصورات وحتى سبب انقراضها.
وكيفية معيشة الديناصورات تكمن جزئيًا فيما إذا كانت من ذوات الدم البارد (مكتسبة الحرارة) كما هو الحال في الزواحف اليوم أو ذوات الدم الحار كما هو الحال في الطيور. فدرجة حرارة جسم ذوات الدم البارد تتغير تبعًا لتغير درجة حرارة الوسط المحيط به. فعلى سبيل المثال، نجد أن درجة حرارة جسم السحلية ترتفع حيث يصبح الهواء أدفأ وإذا برد الهواء فإن السحلية تفقد حرارة جسمها. أما حيوانات الدم الحار فهي ذات درجة حرارة جسم دافئ معتدلة الثبات. ومثل هذه الحيوانات غالبًا ما تكون أكثر نشاطًا من ذوات الحرارة المتغيرة.
ولم يتفق العلماء حول ما إذا كانت الديناصورات من ذوات الدم البارد أو من ذوات الدم الحار. وتقليديًا، اعتُبرت الديناصورات من ذوات الدم البارد، إلا أن العديد من العلماء اليوم يعتقدون أنها كانت قطعًا من ذوات الدم الحار للحفاظ على ارتفاع مستوى نشاطها. ومع هذا، فإن خبراء آخرين يشيرون إلى أن الحيوانات الضخمة تفقد حرارة أجسامها بصورة بطيئة جدًا. ولذلك فإن الديناصورات حتمًا كانت تمتلك درجة حرارة دافئة وثابتة في أجسامها وكانت معتدلة النشاط حتى ولو كانت من ذوات الدم البارد.
الانتشار والموئل
الانتشار الجغرافي

شهد العالم خلال فترة الحقبة الوسطى التي عاشت أثناءها الديناصورات لمُدة 165 مليون عام الكثير من التغيرات على الكوكب، من أبرزها انجراف مواقع القارات وتغير المناخ[45] الذين أثّرا إلى حد كبير على حياة الديناصورات وبيئاتها.[46] لكن هذه الزواحف قطنت عموماً خلال فترة وُجودها على الأرض القارات السبع جمعاءً بما في ذلك أنتاركتيكا وأستراليا وجميع القارات الأخرى.[47][48]
مع مطلع العصر الثلاثي كانت لا تغطي الأرض سوى قارة واحدة عملاقة اسمها بانجيا،[49] وكان مناخ العالم آنذاك أكثر حرارة بقليل مما هو اليوم،[47] وقد كان لربط القارات هذا أهمية بالغة في إتاحة تنقل الديناصورات وانتشارها بين القارات. كانت أولى الديناصورات الحقيقية المَعروفة هي الإيورابتور والهيريراصور الذين عثرَ على بقاياهما في الأرجنتين في طبقات العصر الثلاثي، ولذلك تعتبر دراسات علمي العلاقات التطورية والجغرافيا الحيوية قارة أمريكا الجنوبية مهد الديناصورات وموطن نشوئها الأول.[46] ونظراً إلى أن كل يابسة الأرض كانت مُتحدة آنذاك في بانجيا، فقد اتسعت مناطق انتشار هذه الزواحف بُسرعة لكي تشمل قارات أمريكا الشمالية وأفريقيا وأوروبا فضلاً عن أمريكا الجنوبية نفسها، وبالرغم من هذا فقد كانت أنواع الديناصورات لا تزال قليلة في ذلك الوقت، ولذا فكان كل نوع منها يَشغل مساحة جغرافية واسعة نسبياً. ويَجعل انتشار الديناصورات الثلاثية الواسع هذا من دراستها وتتبع تنوعها وانتشارها أمراً صعباً.[45][46]

مع مجيء العصر الجوراسي المبكر (200 - 170 مليون سنة خلت) كانت لا تزال قارة بانجيا مُترابطة إلى حد ما، وظلت روابط اليابسة قائمة بين كافة القارات السبع، ولذلك احتفظت الديناصورات والفقاريات الأرضية الأخرى بمُعدل توطنها الفريد الذي كان مُنخفضاً بشكل غير اعتيادي مع اتصال العالم ببعضه. وخلال هذه الفترة (العصر الجوراسي) بدأت الديناصورات تهيمن وتفرض سيطرتها الحقيقية على أشكال الحياة الأرضية، وكذلك زادت أكثر من انتشارها الجغرافي عبر العالم.[46] وخلال هذا العصر قبل 180 مليون سنة تقريباً بدأت قارة بانجيا بالتفكك،[50] مما سبب الكثير من التغيرات الجغرافية والمناخية على الكوكب. فخلال تلك الفترة بدأ جليد القطبين بالذوبان وأخذ مستوى البحر[46] بالارتفاع وبغمر مساحات ضخمة من أراضي الكوكب، وقد زاد هذا بدوره من مُستوى الرطوبة ومن ثم الأمطار، فأصبح المُناخ أكثر اعتدالاً بينما نمت غابات غنية بأشكال الحياة المُتنوعة، وساعدت هذه الظروف البيئية الديناصورات على زيادة أعدادها وهيمنتها. كما أخذت القارات بالتفكك والانعزال عن بعضها سريعاً في هذا الوقت، ففي العصر الجوراسي المتأخر انجرفت أمريكا الشمالية شمالاً مُنفصلة بذلك عن أمريكا الجنوبية، وبذلك شكلت أمريكا الشمالية جنباً إلى جنب مع أوروبا جزءاً من قارة لوراسيا، بينما ظلت الجنوبية مُتحدة مع أنتاركتيكا وشبه القارة الهندية وأستراليا مُشكلين غوندوانا قبل أن تنجرف أستراليا بعيداً مُبتدئة بذلك انعزالها الطويل المُستمر إلى يومنا هذا.[51] ومع هذا التفكك السريع وانعزال أجزاء العالم عن بعضها البعض بدأت الديناصورات تخسر قدرتها على التنقل بين القارات، وبدأت معدلات التوطن عند أنواعها بالارتفاع، بينما أخذت ديناصورات كل قارة تتطور وحدها مُنعزلة عن ديناصورات القارات الأخرى.[46]
لا يُوجد أي دليل صريح حتى الآن على حدوث ارتفاع شديد في الحرارة أو تغير ذو قيمة في المناخ خلال العصر الطباشيري، وهو آخر عصور الحقبة الوسطى التي عاشت خلالها الديناصورات. أما بالنسبة للقارات فقد ابتعدت عن بعضها البعض كما لم تبتعد قبلاً سوى مرات قليلة في تاريخ الأرض كله، فقد كانت أنحاء العالم مَعزولة عن بعضها كثيراً وأخذ مُعدل التوطن عند الديناصورات بالارتفاع طوال العصر الطباشيري المتأخر.[46] وخلال هذا العصر قسمت العالم العديد من البحار الضحلة هنا وهناك، ففصل بين غرب أمريكا وشرقها بحر ضحل، وقد كان يَتصل غربها قبل ذلك مع آسيا عبر جسر يابسة قبل أن يَفصل بين نصفي القارة هذا البحر فيُصبح غربها جزيرة معزولة بينما يتحد شرقها مع أوروبا وغرينلاند، أما بقية أراضي العالم الجنوبية فقد تابعت الانجراف مُبتعدة عن بعضها.[52] وكنتيجة لهذا فقد اختلفت أنواع الديناصورات بين القارات، فسادت بشكل عام خلال هذه الفترة في نصف الأرض الجنوبي مجموعات الديناصورات القديمة التي لطالما قطنته، مثل الصوروبودات والأورنيثوبودات والأنكيلوصورات، بينما ظهرت في النصف الشمالي أنواع جديدة تماماً مثل السيراتوبيات والهادروصوريات. وقد ازداد تنوع الديناصورات إلى حد كبير وملحوظ[46] خلال هذا العصر، بينما تقلصت نسبياً المساحة الجغرافية التي يَتوزع عليها كل نوع مع ازدياد هذا التنوع. واستمرت الديناصورات على هذه الوتيرة خلال العصر الطباشيري وتنوعها يَزداد مع ازدياد توطنها[45][52] قبل أن تنقرض تماماً في نهاية ذلك العصر ضمن انقراض العصر الطباشيري-الثلاثي.[53]
البيئات الطبيعية

بالنسبة للبيئات التي قطنتها الديناصورات خلال فترة وُجودها على الأرض بعيداً عن انتشارها الجغرافي فتحديدها أصعب قليلاً. فبشكل عام، من الصعب جداً تخمين طبيعة البيئات التي كانت مَوجودة الماضي باستخدام معلومات الأحافير والصخور التي تكون مُتوافرة عادة.[54] والسبب وراء هذا أنه لا يُعثر سوى على القليل من أحافير الحيوانات أو النباتات التي عاصرت وُجود ديناصور ما (في حال عثرَ عليها أساساً)، علماً بأن دراسة النباتات القديمة بالإضافة إلى دراسة النظائر الجيوكيميائية التي يُمكن من خلالها تحديد الحرارة في تلك العُصور هما الوسيلتان الأساسيتان لمعرفة طبيعة البيئات القديمة (بينما لا تخدم عظام الديناصور سوى في معرفة موقع موته)، وحتى في الحالات التي يُعثر فيها على بقايا تلك النباتات فإن تحديد ما بدت عليه البيئة التي قطنها الديناصور وما كانت معالمها وتضاريسها هو أمر بالغ الصعوبة، مما يَدفع مُعظم الإحاثيين إلى عدم التطرق لهذا الأمر وترك دراسته.[54][55] من المَشاكل الأخرى أيضاً التي تمنع معرفة طبيعة البيئات القديمة بشكل دقيق اختلاف مدى إمكانية حفظ بيئة ما عن أخرى، فبعضها يُمكنها أن تحفظ بشكل جيد بينما تختفي أخرى تماماً بحيث تتعذر معرفتها، وعموماً مُعظم آثار الغابات وأراضي الأنهار المُرتفعة والسهول من المواد العضوية تتحلل وتتآكل ثم تختفي سريعاً، ولذا فإن بقاياها نادراً ما تحفظ بحيث تتيح التعرف عليها. بينما لا يُمكن للعلماء غالباً التعرف إلا على الأراضي النهرية المُنخفضة والسهول الفيضية والدلتا وأراضي البحيرات وما إلى ذلك من البيئات المائية.[55] لكن لا زالت - بالرغم من كل ذلك - توجد بعض الدراسات التي يُمكنها أن تعطي فكرة ما عن طبيعة بيئات هذه الزواحف وأن تلقي الضوء عليها.[56] وعموماً، ربما تكون الديناصورات قد قطنت مجموعة مُتنوعة جداً من البيئات تتراوح من الغابات الرطبة إلى الصحارى القاحلة.[57] وفيما يلي سرد لم يُتوقع أن يَكون البيئات التي عاشت فيها الديناصورات.

نظراً إلى أن كافة أراضي العالم كانت مُتحدة في قارة بانجيا العظيمة بالغة الضخامة خلال العصر الثلاثي، فقد كان من الطبيعي وُجود امتدادات داخلية ضخمة للقارة بعيدة جداً عن أي بحر أو مُحيط، وكنتيجة لهذا فقد ظلت مُعظم المساحات الداخلية للقارة جافة وبعيدة عن الماء وامتدت عبرها صحارى هائلة، بينما لم تكن الرطوبة كافية لنمو النباتات سوى على حواف القارة قرب المسطحات المائية المُحيطية.[58] وقد كانت الحياة مُختلفة جداً في ذلك العصر مع الجفاف السائد، فلم تكن توجد أي نباتات مزهرة أو عشب على الأرض بينما انتشر الصنوبر والسراخس عبر القارة. ومن أسباب الجفاف وقلة التنوع الأحيائي الذين سادا هذا العصر أنه أتى مُباشرة بعد أضخم انقراض في تاريخ الأرض، ولهذا السبب فحتى عندما جاء العصر الثلاثي المتأخر كانت لا تزال الأرض تتعافى من آثار الانقراض.[59] ولذا فقد عاشت الديناصورات خلال هذه الفترة في بيئة جافة ولم يَكن تنوعها كبيراً بعد،[46] وقد تركز وُجودها على ضفاف الأنهار التي كستها السراخس، ومن الأنواع التي قطنت هذه الأراضي ديناصور الهيريراصور المُبكر. أما في المناطق الأبعد من ذلك عن البحار فقد امتدت أراضٍ شبه صحراوية استطاعت أن تنمو فيها بعض أنواع النباتات القصيرة والصغيرة المتكيفة مع المناخ القاحل (وربما بدت تلك المناطق شبيهة بأراضي جنوب أفريقيا اليوم)، ومن الأنواع التي قطنت هذه الأراضي ديناصور البلاتيوصور العاشب جنباً إلى جنب مع الإيورابتور والسينتاصوس.[59]

بحلول العصر الجوراسي بدأت طبيعة الأرض الجغرافية بالتغير مما كان له أثر كبير على بيئات الديناصورات الطبيعية. فقد أخذت قارة بانجيا العظيمة بالتفكك والانقسام مُبتعدة بعضها عن بعض، ومع تكون الصدوع بين القارات بدأت المياه تملأ هذه الأودية والأراضي المُنخفضة المختلفة مُحولة إياها إلى مُحيطات جديدة تفصل بين القارات بعد أن كانت كل المياه تحيط بقارة واحدة فقط (فتكون بذلك المحيط الأطلسي إلى جانب عدة مُحيطات أخرى)، وقد ساعد هذا كثيراً على وُصول المياه إلى قلب بانجيا لكي تسقي صحاراها. فبعد أن دخلت المياه عُمق القارة بدأت الرياح المُصطدمة بالبحار الداخلية بحمل الرطوبة معها إلى الصحارى الجافة،[52][60] فأخذت النباتات تنمو وتزدهر في أنحاء القارات ماحية بذلك الصحارى الثلاثية القديمة ومُكونة مصدراً هاماً جداً لتغذية الديناصورات التي وصلت إلى قمة ازدهارها في هذا العصر، فبعد أن ظلت صغيرة الحجم طوال العصر الثلاثي نمت فجأة لتصبح من أكبر حيوانات الكوكب وأكثرها نجاحاً.[46][60] لكن كما كانت الحال في العصر الثلاثي فقد تركزت الخضرة والديناصورات المختلفة بجانب ضفاف الأنهار، ومع هطول الأمطار نمت غابات من السراخس والغينكجو، ومن الديناصورات التي قطنت هذه الغابات النهرية والسرخسية الستيغوصور. وقد تميز العصر الجوراسي أيضاً بالغابات الصنوبرية الكثيفة التي غطّت القارات خلاله وتألفت من أنواع بدائية من الصنوبر، وقد قطنت هذه الغابات أنواع من الصوربودات مثل المامينكيصور.[59]

ابتعدت القارات عن بعضها كثيراً خلال العصر الطباشيري وبدأت خريطة العالم بأخذ شكل غير بعيد كثيراً عن شكلها الحالي، بالرغم من تغطية البحار الضحلة لجزء كبير منها،[52] وقد كان لهذا أيضاً تأثير على بيئة الكوكب. فوجود أراض بعيدة جداً عن بعضها وفي أجزاء مُختلفة من الكوكب يَعني أيضاً وجود أنواع متعددة من المناخ ومن ثم البيئات الطبيعية.[55] وأيضاً، توجد دلائل على أن الديناصورات قطنت خلال هذا العصر بعض البيئات القطبية اعتماداً على دلائل من موقعين في قارة أستراليا التي كانت لا تزال ضمن الدائرة القطبية الجنوبية في العصر الطباشيري، ويُمكن معرفة ذلك من دراسة أحافير النباتات وبنى الصخور التي كانت متجمدة في الأرض القطبية قبل ملايين السنين.[61]
خلال العصر الطباشيري، انتشرت على أطراف القارات أراضي المستنقعات ودلتا الأنهار - وهي من نوع البيئات التي تحفظ جيداً -، وقد سادت في هذه المستنقعات الأشجار التي تحتاج إلى الماء بشدة مثل السرو. أما من ناحية الديناصورات، فقد كانت هذه البيئات مثالية للديناصورات صائدة السمك مثل السبينوصور. وإلى جانب هذه النوعية من البيئات شبه المائية، فقد كان النوع الآخر السائد من البيئات في العصر الطباشيري هو "الغابات المختلطة" والتي كانت نتيجة بدء ظهور النباتات المزهرة في العصر الطباشيري، وقد كانت هذه الغابات مأوى للعديد من الديناصورات العاشبة مثل الكوريثوصور،[58] كما مثلت في المناطق القريبة من الأنهار موطناً لبعض اللواحم الشهيرة مثل التيرانوصور.[62] من جهة أخرى كان للديناصورات وُجود في بعض البيئات الأكثر اختلافاً خلال هذا العصر، فقد قطنت بعض أنوعها على سبيل المثال في هذه الفترة المناطق القطبية المتجمدة في ألاسكا وأستراليا، ولو أنها لم تكن مختلفة كثيراً عن الديناصورات التي قطنت المناطق المجاورة الأكثر دفئاً.[61] وبالرغم من أن الأحافير لا تقدم الكثير من المعلومات عن بيئات الجبال القديمة، إلا أنه يُعتقد أن بعض أنواعها كانت تقطن هذه المناطق مثل الإدمونتيا. كما أن عدداً كبيراً من الديناصورات الطباشيرية قطنت مناطق الحجر الرملي في الصحارى، ومن هذه الأنواع الغاليميموس[58] والأوفيرابتور والفيلوسيرابتور والبروتوسيراتوبس.
الحجم
تعتبر الديناصورات كمجموعة، إحدى أضخم الكائنات الحية التي سارت على سطح الأرض قديمًا وحديثًا، وقد بلغت بعض الفصائل والأنواع أحجامًا ضخمة جدًا، ومن أبرز الأمثلة على ذلك: طويلات العنق أو الصوروبودات. كان أصغر طويلات العنق حجمًا لا يزال يفوق جميع الأنواع الأخرى التي شاطرته موطنه معظم عهد الديناصورات، أما أكبرها قدًا فكان يفوق جميع الكائنات الحية التي عاشت على الأرض سواءً في الدهور القديمة أو خلال الدهر الحالي، حيث لم تستطع أي دابّة أخرى أن تتفوق عليها في الحجم حتى أثناء العصر الذهبي للثدييات، عندما بلغت الأخيرة أحجامًا ضخمة، فالبالوشيثيريوم أو شبه الأقرن، وهو أضخم الثدييات البريّة البائدة، لا يقترب حتى من مقارعة أكبر طويلات العنق، وكذلك الحال بالنسبة للماموث الكولومبي، أحد أضخم أنواع الفيلة المنقرضة. وحدها بعض الثدييات البحرية المعاصرة تقارب في حجمها حجم طويلات العنق العملاقة أو تتفق عليها، ومنها الحوت الأزرق الذي يصل وزنه إلى 173000 كيلوگرام (381000 رطل) ويزيد طوله عن 30 مترًا (100 قدم).[63] يقترح العلماء عددًا من الإيجابيات كان الحجم الضخم يحققها لطويلات العنق ومنها: الحماية من الضوراي، الاقتصاد في استهلاك الطاقة اللازمة للبقاء، وإطالة أمد الحياة، لكن يظهر أن أهم تلك الحسنات كانت المتعلقة بالغذاء، فالجهاز الهضمي عند الحيوانات الكبيرة أكثر كفاءةً في هضم الطعام من ذاك الخاص بالحيوانات الصغيرة، بما أن الطعام يمضي فترة أطول داخله، وهذا بدوره يجعل من تلك الحيوانات قادرة على الاقتيات على أنواع من الطعام أقل غذاءً لا تستطيع الحيوانات الصغيرة البقاء بمجرد الاقتيات عليها. عُثر على أحافير طويلات العنق، وما زال يُعثر عليها، في طبقات من الصخور ترجع لفترات يُحتمل أن تكون جافة أو موسميّة الجفاف، لذا فإن هذا يدعم ما قال به العلماء من أنها قادرة على الاقتيات على نباتات قليلة الغذاء بفضل حجمها الضخم، لا سيما وأن هذا سيساعدها على البقاء في هكذا بيئة غير ثابتة المناخ.[64]

كانت معظم الديناصورات أصغر حجمًا بأشواط من طويلات العنق، وتشير الأدلّة الحالية أن حجم الديناصورات اختلف من العصر الثلاثي حتى أوائل الجوراسي وأواخره، ثم خلال الطباشيري.[29] يتبيّن عند تصنيف اللواحم الثيروبوديّة إلى فئات وفق قيمة أسيّة، أن معظمها يحتل مركزًا ضمن القائمة التي تحوي نظيرتها التي يتراوح وزنها بين 100 و 1000 كيلوغرام (بين 220 و 2200 رطل)، وبالمقابل ظهر أن اللواحم المعاصرة تقبع بأغلبها ضمن الفئة التي يتراوح وزنها بين 10 و 100 كيلوغرامات (بين 22 و 220 رطلاً).[65] تراوح منوال كتلة جسد الديناصورات بين طن متري واحد وعشرة أطنان متريّة،[66] وهذا يتباين بشكل كبير مع ذاك الخاص بثدييات الدهر الحديث، الذي يترواح بين 2 إلى 5 كيلوغرامات (بين 5 و 10 أرطال) وفق إحصائيات الخبراء في المتحف الوطني للتاريخ الطبيعي في مدينة واشنطن بالولايات المتحدة.[67]
الأكبر والأصغر
.png&filetimestamp=20100101235932&)
خلافًا لما يعتقده البعض من غير المختصين، فإن نسبة ضئيلة فقط من الحيوانات تتحجر هياكلها العظمية وتتحول لمستحاثات مدفونة في باطن الأرض، لذا فإن العثور على أي أحفور أو مستحاثة لأي كائن بائد يُمثل تحديًا لعلماء الآثار. لا يستخرج العلماء هياكلاً عظمية كاملة إلا فيما ندر، ومعظم الأحيان لا يمكن العثور إلا على بضعة عظام متفرقة أو أجزاء من الهيكل العظمي، كذلك فإن طبعات الجلد أو الأنسجة الحيوية المحفوظة أشد ندرة من العظام. يلجأ العلماء إلى مقارنة حجم العظام المعثور عليها حديثًا بالإضافة إلى شكلها بعظام تماثلها في الشكل والحجم حتى يتمكنوا من رسم هيئة الكائن أثناء حياته، إلا أن هذه العملية غير دقيقة على الإطلاق، وهي تبقى مجرّد تخمينات أناس خبراء، ولهذا فإنه من المحتمل ألا يتأكد العلماء على الإطلاق أي الديناصورات كان الأضخم وأيها كان الأصغر.

إن أضخم وأثقل الديناصورات المعروفة ذات الهياكل العظمية الأكثر حفظًا هو الديناصور الزرافي العملاق جيرافاتايتن. اكتُشفت بقايا هذه الديناصورات في تنزانيا خلال الفترة الممتدة بين عاميّ 1907 و1912، وقد جُمعت عظام عدّة أفراد صغيرة الحجم منها في هيكل عظمي واحد يُعرض الآن في متحف همبولت في برلين بألمانيا؛[68] ويصل ارتفاع هذا الهيكل إلى 12 مترًا (39 قدمًا)، ويبلغ طوله 22.5 أمتار (74 قدمًا)، ويُقدّر وزنه لو أضيفت إليه جميع الأعضاء الداخلية والأنسجة الحيوية بما بين 30000 و 60000 كيلوغرام (بين 70000 و 130000 رطلاً). أما أطول الديناصورات فهو الديناصور الطويل ديبلودوكس الذي اكتشفت بقاياه في ولاية وايومينغ بالولايات المتحدة، وجُمع وعُرض في متحف كارنجي للتاريخ الطبيعي في مدينة بيتسبرغ سنة 1907.


كان هناك بعض الديناصورات الأضخم حجمًا من النوعين سالفا الذكر، إلا أن ما يُعرف بشأنها مُشتق من دراسة بعض العظام المتحجرة فقط. اكتشفت معظم العواشب العملاقة خلال عقد السبيعينيات من القرن العشرين أو ما تلاه من عقود، وهذه الديناصورات تشمل: ديناصور الأرجنتين أرجنتينوصور الذي يُعتقد أن وزنه تراوح بين 80000 و 100000 كيلوغرام،[64] والديناصور الخارق سوبرصور الذي يُقدر طوله بحوالي 33 مترًا (110 أقدام)، وديناصور إله الزلازل صوروبوسيدون الذي يظهر بأنه كان أطول الديناصورات قامةً، حيث وصل طوله إلى 18 مترًا (59 قدمًا)،[69] أما أطول الديناصورات بلا منازع فيبدو أنه الديناصور ثنائي التقعّر آمفيوكولياس، الذي يُعرف عن طريق قوس فقريّة واحدة فقط وُصفت عام 1878 وهي الآن مفقودة. يُحتمل أن هذا الديناصور وصل طوله إلى 58 مترًا (190 قدمًا) وفاق وزنه 120000 كيلوغرام (260000 رطل).[64] كان الديناصور الفقري سبينوصور أكبر اللواحم بلا منازع حيث تراوح طوله بين 16 و 18 مترًا (بين 50 و 60 قدمًا)، ووصل وزنه إلى 8150 كيلوغرام (18000 رطل).[70] ومن اللواحم الضخمة الأخرى: ديناصور الريح الجنوبية العملاق جيجناتوصور، الديناصور حاد الأسنان كاركارادونتوصور، وملك الزواحف الطاغية تيرانوصور.[71]

كانت أصغر الديناصورات، باستثناء الطيور المعاصرة، تصل إلى حجم الحمام المستأنس.[72] ومن الديناصورات الصغيرة المميزة: الديناصور شبه الطائر أنتشيورنيس والديناصور المتباهي إيبيدكسيبتركس، الذان لا يفوق طول هيكلهما العظمي 35 سنتيمترًا (1.1 قدم).[72][73] يُعتبر الأنتشيورنيس أصغر الديناصورات حجمًا حاليًا بعد أن وُصفت عينة تعود لفرد بالغ قُدّر وزنه بحوالي 110 غرامات.[73] تشمل قائمة أصغر الديناصورات العاشبة: الديناصور صغير القرون ميكروسيراتوس، وديناصور ونان ونّانوصور، حيث يصل طول كل منهما إلى حوالي 60 سنتيمترًا (قدمين) فقط.[74][75]
النظام الغذائي
هناك اعتقاد بأن السوروبودس ربما كان يخوض في البحيرات والمستنقعات الضحلة لكي يأكل النباتات المائية، أو ربما أكلت أوراق الأشجار كما عملت ديناصورات الدَّكبيل، أو تغذت على النباتات القصيرة التي نمت على خطوط الشواطئ أو السهول المنبسطة كما فعلت الانكيلوسورس والسيراتوبسيان والسيجوسورس.
أما الألوسورس والتيرانوسورس وبعض الثيروبود الكبيرة فمن المحتمل أنها كانت حيوانات صيادة حيث تفترس بشكل أساسي الديناصورات الضخمة آكلة النبات.وربما كانت هذه الديناصورات العملاقة آكلة اللحوم كالثيروبود تأكل كل ما تصادفه من حيوانات ميتة.كما أن بعض الثيروبود الصغيرة كانت تأكل الحشرات أو البيض، وكان بعضها الآخر يصيد الثدييات أو الديناصورات الصغيرة أو الزواحف. ومن الممكن أن بعض الثيروبود الصغيرة هذه كانت نشطة جدًا ويمكنها الجري بسرعة. وربما كان بعضها الآخر مثل الدينونيكس المفترس يصطاد بطريقة جماعية كما تفعل الذئاب والكلاب البرية اليوم.
الحماية من الأعداء
تميزت الديناصورات آكلة النباتات بالعديد من التشكيلات لتحمي نفسها من الثيروبود. ويعتقد أن حجم الثيروبود الضخم قد حماها من معظم الحيوانات المفترسة الأخرى. فالأنكيلوسورس كانت له صفائح عظمية للحماية، والسيراتوبسيان والستيجوسورس ربما قامت باستعمال قرونها وأشواكها لمحاربة الأعداء. كما أن ديناصورات الدَّكبيل كانت تستطيع السباحة في المياه العميقة لتتجنب مهاجمة الثيروبود لها.
السلوك

تُبنى تخمينات سلوك الديناصورات ونمط حياتها عادة على أحافير وموائل ومحاكات حاسوب ميكانيكيتها الحيوية، ثم بمقارنة نتائج هذا مع سلوك الحيوانات الحديثة التي تعيش اليوم أنماطاً حياتية مُشابهة للديناصورات. وبهذا فالفهم الحالي لسلوك الديناصورات يَعتمد إلى حد كبير على التخمين، ولذا فمن المُتوقع أن يَظل موضوعاً مُثيراً للجدل خلال المستقبل القريب. لكن بالرغم من ذلك، فيُوجد اتفاق عام في المُتجمع العلميّ على أن بعض أنواع السلوك الشائعة اليوم بين الطيور والتماسيح - أقارب الديناصورات الحديثة - كانت شائعة أيضاً بين الديناصورات.
اكتشف أول دليل مُحتمل على عيش الديناصورات في قطعان عام 1878، عندما عُثرَ على 31 إغوانودون اعتقد لاحقاً أنهم هلكوا معاً بيرنيسارت - بلجيكا بعد أن سقطوا في شق أرضي عميق ومغمور بالماء.[76] وفي الفترة التي تبعت ذلك اكتشفت عدة مقابر جماعية أخرى لديناصورات. وتشير هذه المَواقع إلى أن الحياة القطيعية كانت شائعة عند العديد من أنواع الديناصورات. وتشير أيضاً مئات أو حتى آلاف آثار أقدام بعض أنواع هذه الزواحف التي عُثرَ عليها حول العالم إلى أن بطيات المنقار (الهادروصورات) ربما تحركت في قطعان عَظيمة معاً، مثلها في ذلك مثل البيسون الأمريكي والظبي القفّاز. تدل آثار بعض الصوربودات على أن هذه الديناصورات كانت تهاجر معاً ضمن مَجموعات تتألف من عدة أنواع مُختلفة، وقد كانت تحدث هذه الهجرات في مَكان واحد على الأقل هو أوكسفوردشاير - إنكلترا،[77] وذلك مع أنه لا يُوجد دليل على وُجود جماعات الديناصورات معاً بحد ذاته.[78] ربما احتشدت الديناصورات في قطعان كبيرة في بعض الأحيان لعدة أغراض بما في ذلك الحصول على الحماية لنفسها ولأطفالها أو للهجرة. وقد عثر على دلائل تشير إلى أن العديد من أنواع الديناصورات - بما في ذلك العديد من الثيرابودات والصوربودات والأنكيلوصورات وذوات القرون والأورنيثوبودات - اجتمعت في بعض الأحيان في مَجموعات تتألف بالكامل من الديناصورات غير البالغة. ومن أمثلة المواقع التي اكتشفت فيها بقايا هذه المَجموعات منغوليا الداخلية التي عُثرَ فيها على أكثر من 20 ديناصور ساينورنيثوميمس، جميعهم تتراوح أعمارهم من سنة إلى 7 سنوات. وقد خمن العلماء أن هذه المجموعة كانت قطيعاً من الديناصورات التي غرقت في الوحل.[79] ذهب تخمين قطعانية الديناصورات هذا إلى أبعد من ذلك أيضاً فوصل إلى اللواحم، فقد ظهرت توقعات بأن بعض أنواع الثيرابودات كانت تعمل معاً في جماعات لصيد الفرائس الكبيرة.[80][81] لكن بالرغم من ذلك، فنمط الحياة هذا ليس مَوجوداً عند أقارب الديناصورات الحديثة (وهي بشكل رئيسي التمساحيات والزواحف الأخرى بالإضافة إلى الطيور) مع أنه توجد بعض الاستثناءات القليلة، وأيضاً يُمكن تفسير الدلائل التحجرية التي تشير إلى أن بعض الثيرابودات كانت تصطاد بشكل جماعي مثل الداينونيكس والألوصور على أنها مُجرد نتيجة للنزاعات الدامية بين الحيوانات على الفريسة، وهو أمر يُرى عند العديد من ضواري ثنائيات الثقوب الحديثة.[82]
برهن اكتشاف الإحاثي جون ر. هورنر عام 1978 لعش ماياصورا في مونتانا على أن الاعتناء الأبوي ظل مَوجوداً لفترة طويلة بين الديناصورات بعد أن ظهر في البداية بين الأورنيثوبودات.[83] عثرَ أيضاً على دلائل تشير إلى أن بعض الديناصورات الطباشيرية الأخرى كانت تعشش معاً في جماعات كبيرة، مثل الصوربودات التايتانوصورية الباتاغونية التي اكتشف عام 1997.[84] اكتشف الأوفيرابتور المنغولي سيتيباتي عام 1993 في موضع تفقيس شبيه بمواضع تفقيس الدجاج، مما يَعني أنه كان مُغطى بطبقة عازلة من الريش كانت تحافظ على حرارة البيض.[85] ولم يَكن هذا الاكتشاف الوَحيد الذي عزز أدلة وُجود الاعتناء الأبوي عند الديناصورات، فمثلاً عُثرَ على بقايا لمجموعة من السيتاكوصورات تتألف من فرد بالغ و34 يَافع عام 2004، وربما يَعود سبب كثرة الحيوانات اليَافعة في هذه الحالة هو كون ذلك المَوقع مكان تعشيش مُشترك للسيتاكوصورات.[86] وبالإضافة إلى ذلك، فقد عُثرَ على جنين ديناصور ماسوسبونديلوس بدون أسنان عام 2005، مما يُشير إلى أن الاعتناء الأبوي كان ضرورياً أحياناً لتغذية الديناصورات اليافعة.[87] تدل أيضاً بعض آثار الأقدام المُكتشفة في جزيرة سكاي شمال غرب أسكتلندا على السلوك الأبوي اتجاه الأطفال عند الأورنيثوبودات.[88] وفضلاً عن كل هذا فقد اكتشفت عموماً بقايا أعشاش وبيوض لمُعظم مَجموعات الديناصورات الكبيرة، ويَبدو أنه من المُرجح أن الديناصورات كانت تتواصل بشكل أو بآخر مع أطفالها بطريقة مُشابهة لطريقة الطيور والتماسيح الحديثة.

ربما كانت أعراف وأهداب بعض الديناصورات - مثل الثيروبودات والهادروصوريات - هشة جداً لكي تستخدم كوسائل دفاع أساسية، ولذا فمن المُرجح أكثر أنها كانت تُستخدم للاستعراض أثناء التزاوج لجذب الإناث أو أثناء المَعارك لإخافة المُهاجمين، ومع ذلك فلا يَعرف العلماء سوى القليل عن التزاوج وتحديد المناطق عند الديناصورات. لكن تشير بعض جراح الرأس التي خلفتها عضات على جماجم ديناصورات إلى أن الثيروبودات - على الأقل - كانت تدخل في معارك ونزاعات عنيفة بين بعضها البعض.[89]
اكتشفت عام 1971 في صحراء غوبي بقايا تعد واحدة من أكثر أحافير الديناصورات نُفساً لدراسة سلوك هذه الحيوانات. فتتضمن هذه البقايا أحافير لفيلوسيرابتورٍ يُهاجم بروتوسيرابتوساً،[90] مما يُعطي للإحاثيين دليلاً على أن الديناصورات كانت بالفعل تهاجم بعضها البعض.[91] ومن الدلائل المُكتشفة الأخرى على مُهاجمة الديناصورات لفرائس حية ذيل إدمونتوصور (وهو ديناصور هادروصوري) معافى جزئياً بعد تأذيه من شيء ما، والطريقة التي تأذى بها الذيل تظهر أن تيرانوصورا عضه لكن الحيوان استطاع النجاة بالرغم من ذلك.[91] أثبت العلماء أيضاً باكتشاف آثار أسنان عُثر عليها في مدغشقر عام 2003 وُجود وحشية (التهام حيوان ما لآخر من نفس نوعه) عند بعض أنواع الديناصورات مثل ثيروبود مادجونغاصور.[92]
بناءً على دلائل الأحافير الحالية من ديناصورات مثل الأوريكتودروميوس، يَبدو أن بعض الأنواع العاشبة من هذه الحيوانات عاشت حياتها جزئياً في جحور تحت الأرض،[93] بينما من المُتحمل أن بعض الأنواع الأخرى الشبيهة بالطيور ربما كانت شجرية (تتسلق الأشجار) خصوصاً الدرومايوصوريات البدائية مثل الميكرورابتور.[94] لكن بالرغم من ذلك، فيَبدو أن مُعظم الديناصورات كانت تعتمد على التحرك فوق الأرض بشكل أساسي. ولذا فإن الفهم الجيد لكيفية تحرك هذه الحيوانات على الأرض هو المفتاح للحصول على نماذج حول سلوكها ونمط حياتها، وبشكل خاص يُعطي علم الميكانيكا الحيوية معلومات بهذا الخصوص. فمثلاً، دراسة القوة التي كانت تبذلها عضلات الديناصورات لتحريك أطرافها وتأثير الجاذبية على بنية هياكلها العظمية يُمكنها أن تعطي فكرة حول السرعة التي استطاعت هذه الزواحف الجري بها،[95] وإذا ما كانت تستطيع الديبلودوكسيات إصدار صوت موجة صدمة بتحريكها لذيلها كمروحة،[96] وإذا ما كانت الصوربودات تستطيع العوم على الماء أم لا.[97]
التواصل

لا تزال طبيعة التواصل بين الديناصورات تُشكل مسألة يكتنفها الغموض، ومجالاً دراسيًا جذّأبًا بالنسبة للعلماء. أقدم عالم الأحياء القديمة الأمريكي "فيل سنتر" في سنة 2008 على دراسة الأدلة المتوافرة المتعلقة بمقدرة حيوانات حُقب الحياة الوسطى، بما فيها الديناصورات، على إصدار الأصوات،[98] فتبين له أنه على العكس مما هو شائع بين الناس، وما تُظهره الأفلام السينمائية، فإن معظم الديناصورات لم تكن قادرة على إصدار أية أصوات على الإطلاق. يُفيد سنتر أنه قام بدراسة توزّع الأعضاء التي تصدر أصواتًا عند كل من الزواحف والطيور، فاكتشف أن الحبال الصوتيّة بداخل الحنجرة يظهر أنها تطوّرت عدّة مرّات عند الزواحف، بما فيها التمساحيات، القادرة على إصدار زئير من حلقها، أما الطيور فهي عديمة الحنجرة، لذا فهي تُصفّر وتغرّد باستخدام عضو آخر يُعرف باسم المصفار، وهذا العضو لا يوجد إلا عند الطيور ولا يرتبط بالحنجرة من قريب أو بعيد، لذا يمكن القول أنه تطوّر بشكل مستقل عن تطور الأعضاء الصوتيّة عند الزواحف. يعتمد المصفار على نظام تشريحيّ مميز عند الطيور يُطلق عليه تسمية "نظام الكيس الهوائي"؛ فإن فقد طائر ما الكيس الهوائي الترقوي الواقع بالقرب من عظم الترقوة، فسيستحيل أبكمًا. يمكن التأكد من وجود هذا الكيس الهوائي عند كائنات منقرضة عن طريق العلامات المميزة أو الفجوات التي يُحدثها للعظام، وبشكل رئيسي الفجوة على العضد، وعند الديناصورات تشير الأدلّة إلى امتلاكها نظم أكياس هوائية معقدة، إلا أن جميع الأجناس والفصائل، عدا جنس واحد، لا يظهر أنها امتلكت كيسًا هوائيًا ترقويًا يُساعدها على إصدار أي صوت ذا شأن. أما الجنس المستثنى فهو جنس الديناصورات هوائيّة العظم (باللاتينية: Aerosteon) التي يظهر أنها طوّرت كيسًا هوائيًا ترقويًا بمعزل عن الطيور لأسباب أخرى لا تتعلق بإصدار الأصوات.[98]
إن أقدم الحيوانات التي تفيد الأدلّة أنها كانت قادرة على إصدار الأصوات باستخدام المصفار هي ما يُعرف بالطيور النقيضة (باللاتينية: Enantiornithes)، ويُحتمل أن أي أركوصور طيريّ آخر سابق لها لم يكن قادرًا على إصدار أي صوت. تفيد الكثير من الأدلّة أن الديناصورات كانت تتواصل بصريًا عوض أن تتواصل سمعيًا، وذلك عن طريق بعض خصائصها الجسديّة المميزة، مثل القرون والأهداب والأعرف وأشرعة الظهر والريش، التي يُحتمل أنها كانت ذات ألوان لامعة تلفت النظر، وفي هذا شبه بسلوك بعض مجموعات الزواحف المعاصرة مثل السحالي، فالكثير من أنواعها صامت تمامًا أغلب الأحيان، لكنه يتواصل مع بني جنسه وغيره من الكائنات باستخدام عروض جسدية تتمحور حول استعمال ألوان جسديّة فاقعة، بل أن البعض مثل الحرابي له القدرة على تغيير لونه وفقًا لحالته النفسيّة، فيكون لونه وهو خائف مختلفًا عن لونه وهو في حالة الاسترخاء.[98]
يقول سنتر وغيره من أنصار نظريته، أنه على الرغم من عدم مقدرة الديناصورات على إصدار الأصوات، فإن بعضها قد يكون لجأ إلى وسائل أخرى لإصدار الصوت بهدف التواصل، فالكثير من الحيوانات المعاصرة بما فيها الزواحف والطيور، تُصدر أصواتًا عديدة لا ملفوظة، منها الهسهسة، صرير الفكين أو الطقطقة بهما، اللجوء إلى ما تقدمه البيئة من حولها كرشّ المياه، أو خفق الجناحين، والأسلوب الأخير يُحتمل أن تكون الديناصورات ذات الأيادي السالبة لجأت إليه كونها كانت ديناصورات مجنحة.[98]
تقترح بعض الدراسات أن الديناصورات بطيّة المنقار العرفيّة أو اللامبيوصوريات كانت تستخدم عرفها الأجوف كغرفة تردد صدى أصواتها المختلفة،[99][100] غير أن سنتر يقول أن هكذا غرف صوتيّة موجودة عند بعض الكائنات المعاصرة، وأن الأخيرة تستخدمها لتفخيم أو تعميق بعض الأصوات غير الملفوظة مثل الهسهسة، ومن أبرز الكائنات عديمة الأوتار الصوتية: الأفاعي، فهي تمتلك غرفًا صوتيةً في جمجمتها تساعدها على تفخيم فحيحها بحال أصبح ما يتهددها من خطر قاب قوسين أو أدنى.[98]
التكاثر


على الرغم من اكتشاف البيض الأحفوري للديناصورات، فإن العلماء لا يعرفون كيف تكاثرت أنواع الديناصورات رغم القول بأن بعضها على الأقل كان يضع البيض كما تفعل معظم الزواحف الأخرى. ومن المحتمل أن تكون الإناث قد بنت أعشاشًا لها في التربة ووضعت فيها عدة بيضات. وربما اعتنت بعض الديناصورات بصغارها بعد فقسها وخروجها من البيض. ومن المحتمل أن بعضها الآخر قد تخلى عن صغاره لتبقى على قيد الحياة بأحسن ما تستطيع.
ولا يملك العلماء إلا أن يخمِّنوا مدى أعمار الديناصورات، ولكن بإمكانهم تحديد الفترة التي استغرقتها الديناصورات لتنمو إلى حجم الاكتمال. ويعتمد معدل النمو على ما إذا كانت الديناصورات من ذوات الدم الحار أو من ذوات الدم البارد، حيث تنمو ذوات الدم الحار بسرعة أكثر من ذوات الدم البارد. فلو كانت الأباتوسورس من ذوات الدم الحار يمكن القول بأنها احتاجت إلى 50 سنة للوصول إلى أوزانها المكتملة التي تقارب 27 طنًا متريًا. ولو كانت الحيوانات من ذوات الدم البارد فلربما احتاجت مائتي سنة أو أكثر لكي تصل إلى نفس الضخامة.
الفسيولوجيا

دار جدال حاد بين العلماء منذ عقد الستينيات من القرن العشرين، وما زال، يتعلّق بما إذا كانت الديناصورات قادرة على تعديل درجة حرارة جسدها بنفسها أم غير قادرة على ذلك. كان العلماء يجمعون في بداية الأمر أن هذه الكائنات لم تكن قادرة على تعديل حرارتها بنفسها على الإطلاق، لذا فقد صنفوها على أنها من ذوات الدم البارد. غير أن بعض الدلائل الجديدة ظهرت مؤخرًا، وهي تدعم الافتراض العكسي، لذا فإن الأغلبية الساحقة من العلماء أصبحوا يوافقون على أن الديناصورات كانت من ذوات الدم الحار، وأصبح الجدال يتركز حول آلياتها الجسدية وتأقلماتها التي خُصصت لتعديل حرارتها.
افترض علماء الأحياء القديمة أن الديناصورات من ذوات الدم البارد عندما اكتشفت أوّل الأحافير، وكان هذا أحد الأسباب التي دفعتهم إلى إطلاق تسمية "السحالي الرهيبة" عليها. جعلت هذه النظرية العلماء يفترضون أن الديناصورات كانت كائنات بطيئة الحركة وبليدة مقارنة مع الزواحف المعاصرة، التي تحتاج إلى مصدر خارجي باعث للحرارة حتى تستطيع رفع درجة حرارتها. ظلّت النظرية القائلة بأن الديناصورات ذات دم بارد نظرية سائدة حتى سنة 1968، عندما نشر عالم الأحياء القديمة الأمريكي، "روبرت ت. باكر"، وهو من الداعمين للنظرية العكسيّة، مقالةً مفصلة تتحدث عن هذا الموضوع وأسباب إيمانه بها، فكان لها كبير الأثر في تغيير واستبدال نظرية الدم البارد التي دامت طيلة سنوات.

تفيد الأدلّة الحديثة أن الديناصورات كانت قادرة على البقاء في المناطق ذات المناخ المعتدل والحرارة الألطف، وهذه المناطق تُعتبر باردةً بالنسبة للكثير من أنواع وفصائل الزواحف والسحالي، لذا يُمكن القول أن بعض أنواع الديناصورات على الأقل كانت قادرة على تعديل درجة حرارة جسدها باستخدام تأقلمات جسدية داخلية. ومن الأدلّة التي تدعم نظرية الدم الحار لدى الديناصورات، اكتشاف ديناصورات قطبيّة في إستراليا والقارة القطبية الجنوبية، وهذه لا شك أنها كانت تعاني من شتاء طويل قارس يدوم ستة أشهر، ولولا استطاعتها التحكم في حرارة جسدها كما الطيور والثدييات لكانت نفقت. أضف إلى ذلك أن أنواعًا كثيرة مريّشة تم اكتشافها، وهذه يُحتمل أن يكون ريشها قد وفّر لها عازلاً حراريًا طبيعيًا، أيضًا أظهرت تحاليل بنية الأوعية الدموية بداخل عظام الديناصورات، أنها مماثلة لتلك الخاصة بذوات الدم الحار. تفيد بنية الهياكل العظمية للواحم الثيروبوديّة وغيرها من الديناصورات، أنها كانت تعيش حياةً نشطة تلائم الأوعية الدموية والقلب الخاص بكائن من ذوات الدم الحار بشكل أفضل من ذوات الدم البارد، بينما تبيّن أن طويلات العنق كان لها خصائص شبيهة بخصائص ذوات الدم البارد أكثر من نقيضتها. يقول العلماء أنه من المؤكد كون بعض الديناصورات حارة الدم وبعضها الآخر بارد الدم، وما زال الجدال قائمًا حول تصنيف الأنواع وفق هاتين الفئتين.[101]
من الأمور التي تزيد الجدال تعقيدًا هو أن خاصيّة الدم الحار تظهر عند الكائنات الحية بسبب أكثر من تأقلم جسديّ واحد فقط. يقوم معظم العلماء بمقارنة الديناصورات بالثدييات والطيور متوسطة الحجم لتفسير تأقلمها الجسدي مع خاصية الدم الحار، حيث يقولون أنها، كما تلك الكائنات المعاصرة، كانت تصرف طاقتها التي تتحصل عليها من الغذاء لرفع درجة حرارة جسدها بما يفوق درجة حرارة محيطها. إلا أن هذه النظرية تعترضها مشكلة أخرى، فإن كانت الديناصورات ستقارن بالحيوانات المعاصرة، فإنها كانت ستعاني من ذات المشاكل أو تمتلك ذات التأقلمات الحرارية التي يفرضها عليها حجمها، فالثدييات والطيور الصغيرة على سبيل المثال تمتلك غطاءً عازلاً للحرارة على جسدها، من شاكلة الفراء والريش، أو الدهون، أما الثدييات الضخمة مثل الفيلة، فتواجه مشكلة الحفاظ على حرارتها بسبب بنيتها الكبيرة، حيث أن بنية الحيوان كلما ازدادت، قلّت نسبة الحرارة التي يلفظها الجلد، الأمر الذي يجبر الحيوان على التأقلم أو البحث عن أساليب أخرى لتخفيض درجة حرارته، وفي حالة الفيلة، فإن البالغة منها تمتلك شعرًا خفيفًا على كافة أنحاء جسدها، وآذانها كبيرة تستخدمها للتهوئة، كما أنها تقوم بالتمرغ بالماء والوحل على الدوام. يُحتمل أن تكون الديناصورات الضخمة قد عانت من ذات المشكلة الأخيرة؛ فحجمها الكبير يفيد بأنها كانت تفقد الحرارة بشكل بطيء، لذا فإنها كانت لتعاني من ارتفاع حرارتها لو لم تمتلك تأقلمًا جسديًا خاصًا، يُرجح بعض العلماء أن يكون مقدرتها على اكتساب الحرارة من محيطها أو فقدانها بدرجة أسرع من الكائنات الأصغر حجمًا، إلا أن هذه النظرية لا تُفسّر كيفيّة تعديل الفصائل العديدة ذات الأحجام الصغيرة حرارة جسدها.
أظهرت التصويرات المقطعية المحوسبة الحديثة للتجويفات الصدريّة الخاصة بالديناصورات وجود ما يُشبه بقايا قلب ذي أربع حجرات شديد الشبه بقلب الطيور والثدييات المعاصرة.[102] تعرّضت هذه النتيجة ومن قال بها إلى انتقاد لاذع في المجتمع العلمي،[103] وذلك بسبب الدراسة التي صنفها كثيرون ضعيفة لا يمكن الاستناد إلى نتائجها، وأنها مجرّد أمنيات لناس خبراء.[104] يرد القائلين بهذه النظرية أنها وإن كانت نتائجها صوريّة، إلا أنه على الرغم من ذلك، فإن التمساحيات والطيور المعاصرة، وهي أقرب الكائنات للديناصورات، تمتلك قلبًا ذي حجرات أربع، لذا لا يُستبعد امتلاك الديناصورات خصائص مماثلة.[105]
أصل الطيور
كان عالم الأحياء البريطاني، طوماس هنري هكسلي، أوّل من افترض أن الطيور هي سليلة الديناصورات المباشرة، وذلك في عام 1868،[106] لكن المجتمع العلمي تخلّى عن هذه الفكرة في أوائل القرن العشرين بعد أن نشر الدكتور وعالم الأحياء القديمة الدنماركي، جيرهارد هيلمان، مؤلفه الذي تحدّث فيه عن أصل الطيور ونشوئها، حيث قال بأنها تتحدر مباشرة من الزواحف غمديّة الأسنان، أسلاف الديناصورات، وكان دليله على ذلك انعدام الترقوة عند الديناصورات ووجودها لدى الطيور.[107] أظهرت الاكتشافات اللاحقة خطأ هذه الافتراضية، حيث عُثر على هياكل عظمية ذات ترقوات، أبرزها هيكل كاسر البيض أوڤيرابتور الذي عُثر عليه سنة 1924،[14] وتبيّن أن العلماء السابقين أخطأوا بين ترقوات كل الهياكل المكتشفة سابقًا واعتقدوا بأنها عظام بين ترقوية.[108] أعاد الدكتور جون أوستروم إحياء نظرية أصل الطيور الديناصوري خلال عقد السبعينيات من القرن العشرين،[109] وقد لاحقت هذه النظرية رواجًا واستحسانًا كبيرًا خلال العقود التالية، خصوصًا بعد التطوّر الكبير الذي طرأ على أساليب تحليل الفروع الحيوية،[110] وبعد اكتشاف الكثير من أنواع الديناصورات الثيروبوديّة الصغيرة المريشة والطيور البدائية،[111] ومن أبرز هذه الاكتشافات: الأحافير من تكوين "تشيان" الصخري في منطقة لياونينغ في الصين، وهي تضم ثيروبودات صغيرة متعددة بالإضافة إلى طيور بدائية.[14] تتشارك الطيور مع الثيروبودات في أكثر مائة خاصية تشريحية، الأمر الذي يدفع العلماء حاليًا لاعتبار الأخيرة أقرب ما يكون من أسلاف الطيور،[112] ويضيفون أن الديناصورات ذات الأيادي السالبة هي الأقرب إليها.[14] اقترحت قلة من العلماء مسارًا تطوريًا آخرًا للطيور، ومنهم من قال بنظرية هيلمان،[113] وقال آخرون أن ذات الأيادي السالبة هي بالفعل أسلاف الطيور، إلا أنها نفسها ليست بديناصورات، بل مجموعة كائنات تطورت تطورًا متوازيًا مع الديناصورات.[114]
عاشت الطيور الحقيقية الأولى في زمن الديناصورات، وكانت مكسوّة بالريش ويصل حجمها لحجم الغراب، وكانت لها أسنان مثل أسنان الزواحف، كما كانت لها أذناب مثل أذناب السحالي، وهذه الأذناب كانت مغطاةً بالريش كذلك الأمر، وقد استدل العلماء على جميع هذه المعطيات من خلال عينة أحفورية تم العثور عليها سنة 1861 في جنوب ألمانيا تمثّل كائنًا شبيهًا بالسحالي إلا أن طبعات ريشه كانت شديدة الوضوح، فقال الخبراء أنه صلة الوصل والحلقة المفقودة بين الزواحف والطيور، وأطلقوا عليه تسمية "الطائر العتيق" (باللاتينية: Archaeopteryx؛ نقحرة: أركيوبتركس). ظهر هذا الاكتشاف بعد سنتين فقط من نشر تشارلز داروين مؤلفه العلمي "أصل الأنواع" (بالإنجليزية: The Origin of Species)، وقد أثار هذا الاكتشاف جدالاً حادًا جديدًا بين العلماء والأشخاص المؤمنين بنظرية التطور وأولئك المؤمنين بنظرية الخلق. إن هذه الكائنات شديدة الشبه بالديناصورات، لدرجة أن إحدى النماذج التي لا تظهر فيها طبعات الريش واضحة، أخطأ العلماء وقامو بتصنيفها على أنها تعود للديناصور مشذب الفك كومبسوگناثوس.[115]
الريش

اكتشف العلماء عددًا كبيرًا من الديناصورات المريشة خلال عقد التسعينيات من القرن العشرين، الأمر الذي دعم النظرية القائلة بأنها أسلاف الطيور بشكل أكبر. وقد عُثر على معظم هذه المستحثات في تكوين "تشيان" الصخري في منطقة لياونينغ، شمالي شرق الصين، التي كانت تُشكل جزءًا من جزيرة قاريّة خلال العصر الطباشيري. يقول العلماء أنه على الرغم من أن الديناصورات المريّشة هي بمعظمها ديناصورات طيريّة، فإنه يُحتمل أن تكون الديناصورات غير الطيريّة أو بعض أنواعها وفصائلها قد اكتسى بالريش. تنقسم الديناصورات المريّشة إلى فئتين: ذوات الريش البدائي، وذوات الريش النصلي، والأولى غير معروفة إلا من فصائل من قاعدة مجموعة العظاءات الحلقية ريشها نحيف ذي بنية خيطية، من شاكلة: الديناصورات مشذبة الفك (باللاتينية: Compsognathidae) وزاحفيّة الجناح الصينية (باللاتينية: Sinosauropteryx)،[116] أما الثانية فهي معروفة من مجموعة ذات الأيادي السالبة وهي تشمل فصائل عديدة مثل كواسر البيض (باللاتينية: Oviraptorosauria) وجارحة الأسنان (باللاتينية: Troodontidae) والطيور المعاصرة.[14][117] لم تنجو نظرية الديناصورات المريشة من الانتقاد اللاذع، فقد قال بعض العلماء أن آثار الريش البدائي ماهي إلا ألياف كولاجينية متحللة كانت تُشكل جزءًا من أهاب تلك الكائنات،[118][119][120] وإن ذات الأيادي السالبة التي تظهر ارتباطًا وثيقًا بالطيور ليست ديناصورات على الإطلاق بل كائنات تطورت بشكل مواز للديناصورات،[114][119] إلا أن هذا الرأي ما زال مرفوضًا من الأغلبية الساحقة من العلماء، لدرجة أن بعضهم يشك في مدى علميّة ما قيل.[121]
الهيكل العظمي
تعتبر الديناصورات المريشة صلة الوصل بين الطيور والزواحف بما أنها تمتلك أبرز سمات الطيور المعاصرة، ألا وهي الريش. إلا أن البنية العظمية لكلا الأصنوفتين هي ما يمثل الدليل الحاسم على هذا الافتراض بالنسبة للعلماء، حيث أن هناك مواقع عديدة في الهياكل العظمية للطيور والديناصورات ذات شبه عظيم، ومنها: العنق، الحوض، المعصم، الذراع، الحزام الصدري، الفريقة، وعظم الصدر أو رافدة القص. يمكن التأكد من مدى وثق العلاقة بين الديناصورات والطيور عند مقارنة هياكلها العظمية من خلال تحليل ودراسة الأفرع الحيوية.
الأعضاء الداخلية

كان للديناصورات اللاحمة الكبرى نظام أكياس هوائية معقد شبيه بذاك الخاص بالطيور المعاصرة، وفقًا لدراسة قام بها الدكتور باتريك أوكونر من جامعة أوهايو، ويفيد الأخير بأنه يُحتمل أن تكون رئات الديناصورات الثيروبوديّة تنفخ الهواء في أكياس فارغة موجودة داخل هيكلها العظمي كما في حالة الطيور، حيث قال: «ما كان يُعتقد سابقًا أنه خاصيّة فريدة بالطيور ظهر بأنه كان موجودًا عند أسلافها القديمة».[122] قام العلماء بوصف نوع جديد من الديناصورات في سنة 2008 أطلقوا عليه تسمية "الديناصور هوائي العظم" (Aerosteon riocoloradensis) وذلك في النشرة العلمية الإلكترونية PLoS ONE، حيث قالوا أن هيكله العظمي يقدم أفضل دليل حتى الآن على وجود ديناصورات ذات أعضاء تنفسيّة شبيهة بالأعضاء التنفسية عند الطيور، بعد أن أظهر التصوير المقطعي المحوسب لهذا الهيكل العظمي وجود أكياس هوائية في تجويفه.[123][124]
من الأدلّة الأخرى التي يستدل بها العلماء على مدى قرابة الديناصورات والطيور، هي استخدام كلا الأصنوفتين لما يُعرف باسم "حصوات القانصة" أو "حصوات الأحشاء"، وهي الحصى التي تبتلعها الحيوانات لتساعدها على الهضم عبر طحن وتكسير الغذاء والألياف القاسية عن طريق تقلبها داخل المعدة، وقد عُثر على عدد من هذه الحصوات داخل بعض الأحافير للديناصورات والطيور القديمة على حد سواء.[125]
الدليل السلوكي
أظهرت إحدى المستحثات المكتشفة حديثًا ديناصورًا من جنس جارحة الأسنان ورأسه تحت يديه، وقد قال الخبراء أن هذه الوضعيّة ماهي إلاّ وضعية النوم،[126] وبالتالي فإن هذا الحيوان بالذات نفق أثناء نومه، ويُلاحظ أن الطيور المعاصرة تنام في كثير من الأحيان ورأسها تحت جناحها، الأمر الذي يُساعدها على الدفء، لذا لعلّ هذه الخاصيّة السلوكيّة إحدى الخصائص التي توارثتها الطيور من أسلافها الديناصورات.
انقراض المجموعات الرئيسية
يعتقد أن الديناصورات قد سيطرت على اليابسة لمدة تقارب 150 مليون سنة، كما سيطرت الزواحف الضخمة الأخرى على السماء والبحر. ومنذ نحو 63 مليون سنة مضت، انقرضت هذه الزواحف الضخمة وسيطرت الثدييات على الأرض.
ولقد وضع الباحثون العديد من النظريات لتفسير اختفاء الديناصورات وبعض الزواحف الضخمة الأخرى. ويُظن أن أقرب هذه التفسيرات نظرية تغيُّر المناخ الأرضي. ففي أواخر العصر الكريتاسي، أصبح الجو باردًا لدرجة لا تحتمله الديناصورات. وكانت الديناصورات كبيرة لدرجة لا يمكن معها أن تسبت في أوكارها، وليس لها ريش أو فرو لحمايتها من البرد. أما الحيوانات الصغيرة فيمكنها السّبات خلال الفترات الباردة. وللثدييات والطيور فرو أو ريش للحماية كما يستطيع بعضها أن يهاجر إلى أماكن أكثر دفئًا لتجنب الطقس البارد. وبهذه السبل، فإن هذه الحيوانات يمكنها النجاة من البرد الذي ربما تسبب في إبادة الديناصورات.
وهناك نظرية أخرى أيضًا تتضمن تغيرات في المناخ. حيث يعتقد بعض الباحثين أن انفجار بعض النجوم القريبة قد أطلق إِشعاعات خطيرة أوجدت جوًا بارداً غير مناسب على الأرض استمر آلاف السنين، فلم تستطع الديناصورات تجنب الإشعاع والبرد وربما كان ذلك سببًا في إبادتها.
ويعتقد بعض الخبراء أن الديناصورات آكلة النباتات لم تتمكن من أكل النباتات الجديدة التي تطورت خلال العصر الكريتاسي مما أدى إلى موتها جوعًا. وبانقراضها انقرضت آكلة اللحوم أيضًا لأنها كانت تتغذى بها.كما يعتقد بعض الخبراء الآخرين أن الديناصورات لم تتمكن من التنافس بنجاح على الغذاء مع الثدييات، لذلك خسرت مقاومتها للبقاء. وهناك نظرية أخرى تقترح أن كوكبًا كبيرًا قد اصطدم بالأرض في نهاية العصر الطباشيري. وطبقًا لهذه النظرية، فإن أثر ذلك كان نشر بلايين الأطنان من الغبار أو بلورات الثلج ـ في حالة اصطدام الكوكب بالمحيط. وقد حجب هذا الحطام ضوء الشمس لمدة تتراوح بين ثلاثة أشهر وستة أشهر. وتمكنت بذور النباتات اليابسة من تحمل هذه الفترة المعتمة إلا أن النباتات نفسها لم تنمُ. ولعدم نمو هذه النباتات، فإن العديد من الديناصورات آكلات النباتات قد ماتت وكذلك الديناصورات التي كانت تتغذى بآكلات النبات. وتسبب الظلام في انخفاض درجة حرارة اليابسة إلى مادون درجة التجمد ولمدة تتراوح بين ستة أشهر واثني عشر شهرًا. وقد أدى التغير في المناخ إلى دمار أكبر في أعداد الديناصورات. ويفترض العلماء بأن الثدييات الصغيرة والطيور قد اتقت من البرد بوساطة فرائها أو ريشها ونجت من الانقراض لأنها كانت تتغذى بالبذور والمكسِّرات والأعشاب المتعفنة. وبعد انحسار الظلام العام، نمت النباتات اليابسة مرة ثانية من البذور والجذور الكامنة.
ويشعر عديد من العلماء بأنه لا يمكن لنظرية واحدة أن تفسر بشكل كامل سبب انقراض الديناصورات، ويقترحون أن الديناصورات وببساطة لم تتمكن من مسايرة التغيرات الحادثة على الأرض مع نهاية العصر الطباشيري. لذلك فإن مزيجًا من الأسباب ربما شاركت في إنهاء عصر الزواحف.
تغيّر المناخ
لم يكن هناك من قلنسوات جليدية عند القطبين خلال ذروة الدهر الوسيط، ويُقدّر العلماء أن مستوى البحر كان أعلى من مستواه الحالي بمقدار يتراوح بين 100 إلى 250 متر (300 إلى 800 قدم)، كذلك كانت درجات الحرارة أكثر تقاربًا من بعضها، حيث كانت الحرارة عند القطبين تختلف عن الحرارة عند خط الاستواء بفارق 25 °مئوية (45 °فهرنهايت)، أما درجات الحرارة في الغلاف الجوي فكانت أعلى من معدلها الحالي بكثير، ففي القطبين على سبيل المثال كانت درجات الحرارة تفوق معدلها الحالي بحوالي 50 °مئوية (90 °فهرنهايت).[127][128]
كانت تركيبة الغلاف الجوي أثناء الدهر الوسيط مختلفة كذلك الأمر عمّا هي عليه اليوم، حيث كانت نسبة ثاني أكسيد الكربون أعلى بإثنا عشر مرة من معدلاتها الحالية، وتراوحت نسبة الأكسجين بين 32% و 35% من إجمالي مكونات الغلاف، مقارنة بنسبة 21% حاليًا. أخذت بيئة كوكب الأرض تتغير بشكل جذري خلال أواخر العصر الطباشيري، فتراجعت نسبة الثورات البركانية بشكل كبير، مما جعل حرارة الأرض تبدأ بالانخفاض تدريجيًا مع انخفاض نسبة ثاني أكسيد الكربون بالجو، كذلك أخذت معدلات الأكسجين تتراجع بشكل ملحوظ، وفي نهاية المطاف انخفضت بشكل عظيم. يفترض بعض العلماء أن التغيّر المناخي بالإضافة إلى انخفاض نسبة الأكسجين أدّى إلى تراجع أعداد الكثير من الأنواع، ولو كان للديناصورات أجهزة تنفسية شبيهة بأجهزة الطيور المعاصرة، فلعلّها واجهت وقتًا عصيبًا خلال هذه الفترة لعدم تمكنها من التنفس بشكل جيّد بفعل تعطش أجسادها الضخمة لكميات كبيرة من الأكسجين.[10]
تأثير الحدث

اقترح البروفيسور والتر ألڤاريز في سنة 1980 نظرية مفادها أن كويكبًا اصطدم بالأرض في أواخر العصر الطباشيري منذ حوالي 65.5 ملايين سنة، وتسبب في اندثار الديناصورات، ويدعم ألڤاريز نظريته بقوله أن طبقات الأرض من تلك الحقبة تُظهر ارتفاعًا مفاجئًا في مستويات الإريديوم.[129] تفيد الأدلّة العديدة التي عُثر عليها أن شهابًا متفجرًا يتراوح قطره بين 5 إلى 15 كيلومترًا (3 إلى 9 أميال) ضرب شبه جزيرة يوكاتان بأمريكا الوسطى، مما أوجد "فوهة شيكسولوب" البالغ قطرها حوالي 180 كيلومتر (110 أميال) وقضى على أشكال متنوعة من الحياة على سطح الكوكب،.[130][131] وما زال العلماء غير واثقين إن كانت الديناصورات في ذلك الحين مزدهرة أم أن أعدادها كانت تتناقص لأسباب طبيعية. يقول بعض العلماء أن اصطدام الكويكب سبب انخفاضًا حادًا غير طبيعي في درجة حرارة الجو، والبعض الآخر يقول العكس، أي أنه تسبب في ظهور موجة حرّ خانقة اجتاحة الكوكب.
يفترض العلماء أن انقراض الديناصورات حصل في وتيرة سريعة للغاية، على الرغم من أن استنتاج ذلك من خلال دراسة الأحافير وحدها مستحيل عمليًا. يقول أنصار هذه النظرية أن الاصطدام تسبب في نفوق الديناصورات بشكل مباشر وغير مباشر على حد سواء، فقد قُتلت الكثير من الحيوانات فورًا بفعل الحرارة الشديدة الناجمة عن الاصطدام، ونفق ما تبقى من الديناصورات بسبب انخفاض حرارة الأرض بعد أن حُجبت أشعة الشمس بفعل الغبار الكثيف والركام الناجم عن الصدمة.
قام علماء أمريكيون بالتعاون مع آخرون تشيكيّون في شهر سبتمبر من سنة 2007 لمحاولة تحديد مصدر الكويكب الذي أوجد فوّهة شيكسولوب، وباستخدام المحاكاة الحاسوبيّة استنتجوا أن احتمالية أن يكون مصدر الكويكب هو الكويكب العملاق المُسمى "المعمداني" (بالإنجليزية: Baptistina) تصل إلى 90%. يصل قطر هذا الكويكب إلى 160 كيلومترًا (100 ميل)، وهو يقع ضمن حزام الكويكبات بين المريخ والمشتري، ولعلّ كأحد الكويكبات الأصغر حجمًا وغير المسماة بعد، والبالغ قطره 55 كيلومترًا (35 ميلاً) اصطدم به منذ حوالي 160 مليون سنة، وقد أدّى هذا الاصطدام إلى تفتيت المعمداني مما أوجد عنقودًا كويكبيًا لا يزال موجودًا حتى اليوم ويُعرف باسم العنقود المعمداني. أظهرت الحسابات أن بعض تلك الفتات توجهت نحو الأرض بعد الاصطدام، وأحدها كان حجرًا نيزكيًا ذي قطر يساوي 10 كيلومترات (6 أميال)، اخترق الغلاف الجوي واصطدم بشبه جزيرة يوكاتان منذ حوالي 65 مليون سنة، وتسبب في وجود فوّهة شيكسولوب.[132]
من التفسيرات المشابهة لانقراض الديناصورات، النظرية الأكثر جدلاً والتي تقول أن عبور النجم الثنائي الافتراضي "نيميسس" سحابة أورط التي يُعتقد أنها تشكل مصدر المذنبات الرئيسي في النظام الشمسي، أدّى إلى تبعثر عدد كبير منها، واصطدام إحداها، أو بعضها حتى، بالأرض في نهاية المطاف مما تسبب بنفوق الكثير من الكائنات الحية.[133] وكما في نظرية الكويكب، فإن حصيلة اصطدام المذنب أو هطول الكثير منها، كانت انخفاض درجات الحرارة بشكل سريع، تلاها فترة طويلة من البرد والصقيع أفنت ما الديناصورات الناجية من الانقراض.[133]
أفخاخ هضبة الدكن

قبل سنة 2000، كان القائلون بنظرية الانقراض بفعل أفخاخ الدكن البازلتيّة الفيضيّة يربطون بينها وبين النظرية القائلة بحدوث الانقراض بشكل تدريجي طيلة قرون عديدة. والأفخاخ الفيضيّة هذه عبارة عن تكوينات حممية بازلتيّة ظهر بعضها نتيجة ثوران بركاني عظيم وبعضها الآخر نتيجة ثورات متعددة، وكوّنت هضاب كثيرة وسلاسل جبليّة على مختلف القارّات، ويقول أنصار هذه النظرية أن الأفخاخ بدأت بالظهور منذ حوالي 68 مليون سنة، واستمرت تظهر طيلة مليونيّ عام. غير أن بعض الدلائل تشير إلى أن ثلثيّ الأفخاخ ظهر خلال مليون سنة فقط، أي منذ حوالي 65.5 ملايين سنة، وبالتالي فإن كانت هذه النظرية صحيحة، فإن هذا يعني أن الأفخاخ سببت أنقراضًا سريعًا للديناصورات وليس انقراضًا تدريجيًا، حيث يُحتمل أن تكون كافّة الأنواع والفصائل قد اندثرت خلال آلاف السنين، لكن هذه الفترة تبقى طويلة على الرغم من ذلك عند مقارنتها بالفترة التي يُفترض أن الديناصورات استغرقتها لتندثر بفعل اصطدام كويكب أو مذنب بالأرض.[134][135]
يُحتمل أن تكون أفخاخ الدكن قد ساهمت بانقراض الديناصورات عبر أكثر من وسيلة، بما في ذلك إطلاقها لكميات كبيرة من الغبار والهباء الكبريتي في الجو، الأمر الذي حجب أشعة الشمس، فلم تعد النباتات قادرة على توليد غذائها عبر عملية التركيب الضوئي، فذبل الكثير منها، ومات الكثير من الديناصورات العاشبة بعد أن فقدت مصدر غذائها، ثم لحقتها اللواحم بدورها. أيضًا يُحتمل أن النشاط البركاني أسفر عنه انبعاثات كثيفة لثاني أكسيد الكربون، مما يعني أن الأرض عانت من احتباس حراري، بعد أن انقشع الغبار والهباء، كان له كبير الأثر على الحياة النباتية والكائنات التي اعتمدت عليها.[135] يقول بعض العلماء أنه قبل الانقراض الجماعي للديناصورات، ساهمت الانبعاثات الغازية البركانيّة خلال تكوّن أفخاخ الدكن، ساهمت "بحدوث احتباس حراري عالميّ كبير، وتشير بعض البيانات إلى أن معدّل ارتفاع درجات الحرارة خلال النصف مليون سنة السابقة على الاصطدام في شيكسولوب وصل إلى 8 °مئوية (14 °فهرنهايت)".[134][135]
قال البروفيسور لويس ألڤاريز (توفي سنة 1988) خلال الفترة التي كان فيها القائلون بنظرية أفخاخ الدكن يربطون بينها وبين الانقراض البطيء للديناصورات، أن علماء الأحياء القديمة أضلّتهم بيانات متفرقة، وأنه لا يمكن القول بصحة رأيهم، لكن هذا لم يؤخذ بعين الاعتبار إلا بعد سنوات من الدراسات الميدانية المكثفة لمواقع الاكتشافات، وفي نهاية المطاف أخذت الأغلبية الساحقة من علماء الأحياء القديمة تتقبل فكرة أن الانقراض الجماعي الذي حصل في أواخر العصر الطباشيري إنما هو نتيجة اصطدام جسم كوني بالأرض على الأغلب. لكن على الرغم من ذلك، يقول البروفيسور والتر ألڤاريز، ابن البروفيسور سالف الذكر، أن الأرض كانت عرضة لتغييرات هائلة طيل الفترة السابقة على الاصطدام، ومن هذه التغييرات: انخفاض مستوى البحر وحصول ثورات بركانية هائلة كوّنت أفخاخ الدكن، ولعلّ هذه الأحداث أيضًا ساهمت في اندثار الديناصورات، وكان الاصطدام بمثابة الضربة القاضية.[136]
فشل التأقلم مع الظروف المتغيرة
قال بعض العلماء أن النباتات المزهرة مغطاة البذور أصبحت تُشكل جزءًا مهمًا من النظام البيئي بحلول منتصف العصر الطباشيري، مستبدلة بذلك عاريات البذور مثل الصنوبريات التي هيمنت على المنظر الطبيعي طيلة ملايين السنين. يُظهر براز الديناصورات المتحجر أن بعض آكلات الأعشاب اقتاتت على مغطاة البذور، بينما استمرت الأغلبية منها تقتات على عاريات البذور بشكل رئيسي، وقد تبيّن من خلال التحاليل الإحصائية أن الديناصورات لم تتفرع لأنواع وفصائل جديدة مختلفة خلال أواخر العصر الطباشيري، كما كان يُعتقد في السابق، فقال داعموا هذه النظرية أن الديناصورات فشلت في التأقلم مع المتغيرات البيئية حولها، ولم تستطع أن تزدهر لتظهر منها أشكال جديدة، وكان هذا الجمود في تاريخ نشوئها هو ما حكم عليها بالفناء.[32]
ناجون محتملون حتى العصر الپاليوسيني
Non-avian dinosaur remains have occasionally been found above the K-Pg boundary. In 2000, Spencer Lucas and colleagues reported the discovery of a single hadrosaur right femur in the San Juan Basin of New Mexico, and described it as evidence of Paleocene dinosaurs. The rock unit in which the bone was discovered has been dated to the early Paleocene epoch, approximately 64.8 million years ago.[137] If the bone was not re-deposited by weathering action, it would provide evidence that some dinosaur populations may have survived at least half a million years into the Cenozoic.[138] Other evidence includes the presence of dinosaur remains in the Hell Creek Formation up to 1.3 m (4.3 ft) above the Cretaceous–Paleogene boundary, representing 40,000 years of elapsed time. This has been used to support the view that the K-Pg extinction was gradual.[139] However, these supposed Paleocene dinosaurs are considered by many other researchers to be reworked, that is, washed out of their original locations and then re-buried in younger sediments.[140][141][142] The age estimates have also been considered unreliable.[143]
تاريخ الدراسة
 مقالة مفصلة: تاريخ علم المتحجرات
مقالة مفصلة: تاريخ علم المتحجرات




لدى العلماء العديد من الطرق لمعرفة الديناصورات. واحدة من أهم هذه الطرق تتم من خلال دراسة أحافير الديناصورات. فعلى سبيل المثال، فإن سن الديناصورات يمكن أن يجعل الخبير يتوقع إن كان الحيوان آكل نبات أم آكل لحوم.
يسمى العلماء الذين يدرسون الأحافير علماء الإحاثة (الأحافير). ويتعلم العلماء عن الديناصورات من خلال ملاحظة الحيوانات التي لها صفات مشابهة لصفات الديناصورات. فعلى سبيل المثال، يمكنهم دراسة الفيلة وكذلك وحيد القرن في الأدغال لمعرفة المزيد عن حياة الحيوانات الكبيرة التي عاشت على الأرض.

اكتشافات الديناصور
لم يكن أحد يعلم بأن الديناصورات قد وُجدت أبدًا واستمر ذلك حتى بداية القرن التاسع عشر الميلادي. حيث إن الناس الذين عثروا على عظمة أو سن لديناصور لم يعرفوا كُنْهها. وفي عام 1822م وجدت زوجة طبيب انجلترا اسمه جدايون مانتل سنًا كبيرة مدفونة بشكل جزئي في الصخر. وأطلعت عليها زوجها الذي كان يجمع الأحافير. وعلم مانتل أن السن تشبه سن سحلية في جنوب أمريكا الجنوبية اسمها الإجوانة. واقترح أن هذه السن آتية من أحد الزواحف الضخمة التي تشبه الإجوانة وسماها الإجوانودون (سن الإجوانة).
وخلال بضع سنين، تم اكتشاف بقايا أنواع أخرى من الزواحف الكبيرة المنقرضة. وفي عام 1841م، اقترح السيد ريتشارد أوينْ وهو عالم إنجليزي أن هذه الزواحف ترجع لمجموعة من الزواحف المختلفة مما نعرفه من حيوانات اليوم. وقد أعطاها أوين اسم ديناصوريا وأصبحت هذه المجموعة تُعرف بالديناصورات ولكن تصنيفها أصبح معقدًا.
وفي أواخر القرن التاسع عشر وبداية القرن العشرين اكتُشفت بقايا كثيرة من الديناصورات في غربي أمريكا الشمالية وأوروبا وآسيا وإفريقيا.
ومن أغنى المناطق في العالم بهذه البقايا الديناصورية منطقة وادي نهر الغزال الأحمر في جنوبي ألبرتا في كندا. وهناك منطقة أخرى غنية جدًا بالبقايا المتحجرة للديناصورات وهي إقليم زنج يانغ في الصين على مشارف صحراء جوبي. كما توجد رواسب وأحافير كبيرة أيضًا في بلجيكا و منغوليا و تنزانيا و ألمانيا ومناطق عديدة أخرى في العالم. أما في أستراليا فلم يكتشف الكثير من بقايا الديناصورات وما تم الكشف عنه أتى بصورة رئيسية من كوينزلاند ومن فكتوريا مؤخرًا. وتنتمي الديناصورات التي تم كشفها في أستراليا إلى الأونيثوبودس والأنكيلوسورس المصفح.
العمل على أحافير الديناصور
تقوم المتاحف والمعاهد والمؤسسات التعليمية بدعم العلماء الباحثين في أحافير الديناصورات ودراستها. ويقوم علماء الإحاثة (الأحافير) بالبحث عن الأحافير في الأماكن التي تعرّت فيها اليابسة بفعل الريح والمياه حتى أصبحت الطبقات الصخرية العميقة الحاملة للأحافير مكشوفة وبعدها يقومون بإزالة الصخور التي تعلوها.
وفي كثير من الحالات، يقومون باستخراج الأجزاء الصخرية المحتوية على الأحافير. ومن ثم تغطية الصخور والأحافير بالقماش والجص الباريسي. ويجف الجص ليصبح على شكل غلاف صلب واق وبعدها يتم شحن الأحافير إلى المختبر.
وفي المختبر، يقوم العاملون بتنظيف العظام والأسنان وإصلاح العظام المكسورة. وبعدها ربما يقوم المختصون بإعادة تركيب الهيكل العظمي بوضع العظام بعضها مع بعض على إطار معدني. وفي بعض الأحيان تستبدل، العظام المفقودة بقطع من الألياف الزجاجية أو الجص أو البلاستيك. ونادراً ما يكتشف العلماء جميع العظام لديناصور ضخم ومن ثم فإنهم يقدرون طول الحيوان اعتمادًا على العظام التي وجدوها.
تقوم بعض المتاحف بعمل نماذج للديناصورات لأجل العرض إذ يدرس الخبير الهيكل العظمي، ويحاول تصور شكل الحيوان بعضلاته وجلده ثم يبنون إطارًا من المعدن يشابه الهيكل العظمي، ويثبتون فيه الأسلاك والأغطية بحيث يتشكل على الهيئة المتصورة لجسم الديناصور، ويغطى أخيرًا بالجلد ويطلى باللون المناسب ليضفي عليه شكلاً حقيقيًا.
"نهضة الديناصورات"

اشتدت موجة التنقيب عن أحافير الديناصورات بشكل بارز منذ عقد السبعينيات من القرن العشرين وما زالت، ويُعزى ذلك إلى اكتشاف الدكتور جون أوستروم للديناصور ذو المخلب الرهيب داينونيكوس، الذي أفادت الأدلّة المتمحضة عن دراسته أنه كان مفترسًا نشطًا حار الدم، أي عكس الفكرة التي كانت سائدة آنذاك، ألا وهي أن الديناصورات كائنات بليدة باردة الدم. تحوّل علم الأحياء الفقارية القديمة إلى علم مهم يُدرّس في الكليّات والجامعات حول العالم بفضل تلك الموجة من الدراسات والتنقيبات، خصوصًا بعد أن وجّه العلماء اهتمامهم نحو مناطق جديدة لم تجر فيها أي تنقيبات في السابق، فاكتُشفت أنواع وفصائل وأجناس جديدة في الهند وأمريكا الجنوبية ومدغشقر والقارة القطبية الجنوبية، إلا أن أهم تلك الاكتشافات وأكثرهًا جذبًا للناس من غير المختصين والعلماء على حد سواء كانت تلك التي برزت في الصين. كذلك أدّى الانتشار الموسع لتطبيق علم الكلاديسيات أو دراسة الأفرع الحيوية، الذي يقوم بتحليل العلاقة بين الكائنات الحية الموجودة والمندثرة، إلى تصنيف الديناصورات بشكل أكثر دقة، مما ساعد على سد الثغور في سجل المستحثات الناقص.
بقايا الأنسجة الحيوية والحمض النووي
كُشف النقاب عن إحدى أفضل المستحثات ذات الأنسجة الحيوية المحفوظة في سنة 1998، وكان هذا الأحفور يعود لفرخ ديناصور مخلب سكيبيو سكيبيونكس عُثر عليه في منطقة "بترارويا" في إيطاليا، وقد ظهرت فيه أجزاء من الأمعاء والقولون والكبد والعضلات والقصبة الهوائية.[43]

أعلنت عالمة الأحياء القديمة الأمريكية، "ماري هيجبي شويتزر"، في عدد شهر مارس لسنة 2005 من مجلة العلوم الأكاديمية، أنها اكتشفت وفريقها موادًا طيّعة تشبه الأنسجة الحيوية الطبيعية بداخل عظمة ساق تيرانوصور تبلغ من العمر 68 مليون سنة، عُثر عليها في التكوينة الصخريّة المعروفة باسم "تكوينة جدول الجحيم" (بالإنجليزية: Hell Creek Formation) في ولاية مونتانا، وقد قام الفريق بتمييه هذه الأنسجة بعد استخراجها لغرض دراستها.[44] بعد أن قام العلماء بتنظيف تلك العظمة طيلة أسابيع عدّة لاستخراج ما علق فيها من تراب ومعادن، تبيّن أن فيها عدد من البنى السليمة، من شاكلة الأوعية الدموية ومصفوفات وألياف عظميّة، كذلك أظهرت الفحوصات الدقيقة تحت المجهر أن هذه الأنسجة الحيوية المفترضة حافظت على بنيتها الدقيقة حتى على المستوى الخليوي. وعلى الرغم من كل هذا، فإن طبيعة وتكوين هذه المواد التي عُثر عليها لا تزال غير واضحة، وما زال العلماء منكبون على دراستها.[44]
قال الدكتور طوماس كاي من جامعة واشنطن بتاريخ 30 يوليو سنة 2008 أن المواد التي عُثر عليها في عظمة ساق التيرانوصور ليست بأنسجة حيوية على الإطلاق بل بيوفيلم تكوّن بعد أن شمّعت البكتيريا الفراغات حيث كانت الأنسجة الحيوية والخلايا تقع.[144] كذلك أفاد الباحثون أن ما اعتقد أنه بقايا خلايا دموية بفعل احتوائه على ذرّات حديديّة، إنما هو في واقع الأمر أجسام كرويّة دقيقة تحمل ذرّات حديد اكتُشفت في الكثير من مستحثات كائنات منقرضة أخرى مثل الأمونيت. وكان وجود هذه الأجسام في بعض أنحاء جسد الأمونيت حيث لا وجود لأوعية دموية مفتاح الاستنتاج الذي توصّل إليه كاي، وهو أنه لا يمكن ربط وجودها بوجود أوعية دموية.[145]
أفاد بعض العلماء مرتين فقط أنهم تمكنوا من استخراج حمض نووي سليم من مستحثات تعود لديناصورات، إلا أنه عند التدقيق بهذه الإدعاءات تبيّن أنها غير موثوقة.[146] إلا أنه تمّ الاستدلال على شكل الببتيد في أحماضها الأمينية عبر تحليل تاريخ نشوئها وتطورها بالاستناد إلى سلسلة مورثات أقرب أقاربها المعاصرة من طيور وزواحف.[147] إضافة إلى ذلك تمّ الإدعاء بتأكيد وجود عدد من البروتينات، بما فيها الهيموجلوبين،[148] في عدد من المستحثات.[149]
على مدى الـ100 عام الماضية، تم العثور على عدد من بيض وأعشاش الديناصورات المتحجرة، ولكن من النادر للغاية العثور على بيضة تحفظ الجنين في حالة جيدة، وفقا لتقرير سكاى نيوز عربية .
وفى دراسة جديدة نُشرت فى دورية "آي سينس" كشفت حقائق مذهلة عن بيضة ديناصور متحجرة عمرها 72 مليون سنة اكتشفت لأول مرة عام 2000 في مدينة غانتشو بمقاطعة جيانغشي جنوب الصين وتوصل الباحثون في دراستهم إلى حقائق مدهشة عن الأفيرابتوروصورات (مجموعة من الديناصورات ذات المنقار وهى وثيقة الصلة بالطيور التى عاشت منذ حوالى 130 مليون إلى 66 مليون سنة، وتشترك فى العديد من السمات مع أقاربها من الطيور الحالية)، حيث اتخذ جنين هذا النوع من الديناصورات وضعية ملتوية مميزة قبل الفقس، وهو سلوك تميزت به الطيور، ويُرجع العلماء هذه الوضعية الغريبة إلى احتمال أن يكون سلوك دفن البيضة قد تطور أولاً بين الديناصورات خلال العصر الطباشيري.
وتقول ويسوم ما من جامعة برمنغهام بالمملكة المتحدة: "معظم أجنة الديناصورات "غير الطيرية" المعروفة غير مكتملة بهياكل عظمية مفككة، حيث لم يتم التعرف من قبل على هذا الوضع في الديناصورات غير الطيرية وعثر على العينة الأحفورية أول الأمر عام 2000 في صخور يقدر عمرها بنحو 70 مليون سنة، لكن عمر الحفرية الدقيق لا يزال غير مؤكد.
ورغم الاشتباه في كون الصخور تحتوي على أحد أحافير البيض، لكن كان مآلها التخزين، وتم نسيان الأمر، وبعد 10 سنوات اكتشف موظفو متحف "ينگليانگ ستون" لتاريخ الطبيعة أثناء فرز الصناديق، ومن هنا تم اكتشاف الحفرية وقالت ليدا شينغ من جامعة الصين لعلوم الأرض في بكين: "تعرف عليها موظفو المتحف على أنها بيض ديناصور ورأوا بعض العظام على المقطع العرضي المكسور لإحدى البيضات"، ثم تم تجهيز الحفريات للدراسة، وكشف النقاب عن الجنين المخبأ بداخل البيضة، والذي وأطلق عليه الباحثون اسم (بيبي ينگليانگ)".
وضعية غريبة وفى الدراسة، أفاد الباحثون أن الجنين وجد ملتفًا حول نفسه بإحكام شديد واضعًا رأسه بين أصابع قدميه، بينما كان الظهر ملتفًا على طول القطب الحادة للبيضة، في وضع لم يكن يعرف من قبل في ديناصور غير طيري، حيث يعرف هذا الوضع بشكل خاص فى مرحلة متأخرة في جنين الطيور الحديثة.
وتقول دارلا زيلينيتسكي الباحثة الرئيسية وعالمة الحفريات بجامعة كالغاري والمتخصصة في بيض الديناصورات: "لم أصدق عيني لأن الجنين كان سليمًا والعينة كانت محفوظة تمامًا".
وأضافت أنها "عينة رائعة، أعمل على بيض الديناصورات منذ 25 عاماً لكني لم أرَ أي شيء مثل هذه العينة قبل الآن، حيث لا يوجد لدينا إلا عدد قليل للغاية من الهياكل العظمية لأجنة الديناصورات ناهيك عن أن تكون مكتملة النمو ومحفوظة بالكامل".
الدراسة أوضحت أن طول البيضة يبلغ نحو 17 سم، ويقدر طول الديناصور بداخلها بحوالي 27 سنتيمتر من الرأس إلى الذيل، في حين يعتقد الباحثون أنه كديناصور بالغ لو عاش لكان طوله حوالي مترين إلى ثلاثة أمتار.
كما تشير مقارنة العينة بأجنة الأفيرابتوروصور الأخرى في المرحلة المتأخرة إلى أن وضعية الجنين تسير إلى مرحلة ما قبل الفقس، وطور الأفيرابتوروصورات أوضاع شبيهة بالطيور في وقت متأخر من فترة الحضانة في الطيور الحديثة، إذ ترتبط هذه الحركات الجنينية الملتوية بنجاح عملية الفقس، ويتحكم في هذا السلوك الجهاز العصبي المركزي.
ويؤكد الباحثون إنهم سيواصلون دراسة هذه العينة النادرة بعمق أكبر، باستخدام تقنيات تصوير مختلفة لتصوير تشريحها الداخلي، مثل عظام الجمجمة، وأجزاء الجسم الأخرى التي لا تزال مغطاة بالصخور ليعرفوا على وجه الدقة منشأ هذا السلوك في مرحلة ما قبل الفقس.[150]
خط زمني للأوصاف التصنيفية لأشهر الديناصورات
 |
التصوير الثقافي


استحوذت الديناصورات على مخيلة الناس وأصبحت تُشكل جزءًا مهمًا من الثقافة الإنسانية منذ اكتشافها، وذلك بسبب الفكرة الشائعة التي سيطرت على أذهان البشر، بأن تلك الكائنات مهيبة الشكل وعملاقة في الحجم. تُستخدم كلمة "ديناصور" في الخطاب العاميّ بين الناس لوصف شيء أو شخص ضخم الجثة أو بطيء الحركة، أو عفا عليه الزمن ويُتوقع زواله أو موته،[151] الأمر الذي يعكس الفكرة السائدة عند الناس التي تقول أن الديناصورات كانت وحوشًا بائدة سيئة التهيؤ غير قادرة على التأقلم مع محيطها ومواجهته.

ظهر تمثيل الديناصورات في الثقافة العامّة لأوّل مرة في إنجلترا خلال العصر الڤيكتوري، عندما عُرضت لها منحوتات في حديقة القصر الكريستالي في لندن سنة 1854، أي بعد 3 عقود من وصف الديناصورات وصفُا علميًا لأول مرة. لاقت هذه المنحوتات شعبيّة كبيرة لدرجة أن التجارة بتماثيل مصغرة تماثلها شكلاً مخصصة للبيع للعامّة كانت إحدى أكثر التجارات ربحًا. افتتحت معارض للديناصورات في الحدائق والمتاحف حول العالم خلال العقود القليلة اللاحقة، مخصصة لتعريف الأجيال اللاحقة على تلك الكائنات بأسلوب مثير.[152] أدّت الشعبية المتزايدة للديناصورات إلى قيام بعض الوجهاء بتمويل بعثات وحملات التنقيب عن بقاياها، الأمر الذي نجم عنه تحقيق عدد من الاكتشافات المهمة، وتبقى حرب العظام إحدى أبرز وجوه ونتائج الشعبية الكبيرة للديناصورات التي دفعت المتاحف إلى التنافس فيما بينها لعرض أكبر عدد من الأحافير للعموم.[153]
كان من الطبيعي ظهور الديناصورات في الروايات الأدبية والأفلام السينمائية وغيرها من الوسائط الإعلامية بعد أن حظيت بشهرة واسعة في صفوف العامّة. ذُكرت الديناصورات لأول مرة في عمل أدبي سنة 1852، وذلك في رواية للكاتب تشارلز ديكنز تحمل عنوان "البيت الكئيب"،[154] ومنذ ذلك الحين والديناصورات تظهر في قصص خيالية عديدة، لعلّ أبرزها كتاب العالم المفقود من سنة 1912، للسير آرثر كونان دويل، وفيلم "كينگ كونگ" الإبداعي من سنة 1933، وگودزيلا من سنة 1954، ورواية الحديقة الجوراسية من سنة 1990 والفيلم المقتبس عنها، وأعمال كثيرة غيرها. استخدمت شركات عديدة الديناصورات في الإعلان عن منتجاتها على شاشة التلفاز، وذلك إما في سبيل تسويق هذه المنتجات، أو لتظهر كيف أن منافساتها ليست قادرة على أن تتفوق عليها، فهي "كالديناصورات": بطيئة وبليدة وبائدة.[155]
انظر أيضاً
- المسار الحيواني
- النظام الغذائي وتغذية الديناصورات
- التاريخ التطوري للحياة
- قائمة تشكيلات الصور المقترنة بالديناصورات
- قائمة الديناصورات
- ديناصور حي
مرئيات
| مقارنة أحجام الديناصورات المختلفة بحجم الإنسان. |
الهوامش والمصادر
- ^ Matthew G. Baron; Megan E. Williams (2018). "A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution". Acta Palaeontologica Polonica. 63. DOI:10.4202/app.00372.2017.
- ^ Andrea Cau (2018). "The assembly of the avian body plan: a 160-million-year long process" (PDF). Bollettino della Società Paleontologica Italiana. 57 (1): 1–25. DOI:10.4435/BSPI.2018.01.
- ^ Ferigolo, Jorge; Langer, Max C. (2007). "A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone". Historical Biology. 19 (1): 23–33. Bibcode:2007HBio...19...23F. doi:10.1080/08912960600845767. ISSN 0891-2963. S2CID 85819339.
- ^ Langer, Max C.; Ferigolo, Jorge (2013). "The Late Triassic dinosauromorph Sacisaurus agudoensis (Caturrita Formation; Rio Grande do Sul, Brazil): anatomy and affinities". Geological Society, London, Special Publications (in الإنجليزية). 379 (1): 353–392. Bibcode:2013GSLSP.379..353L. doi:10.1144/SP379.16. ISSN 0305-8719. S2CID 131414332.
- ^ Cabreira, S.F.; Kellner, A.W.A.; Dias-da-Silva, S.; da Silva, L.R.; Bronzati, M.; de Almeida Marsola, J.C.; Müller, R.T.; de Souza Bittencourt, J.; Batista, B.J.; Raugust, T.; Carrilho, R.; Brodt, A.; Langer, M.C. (2016). "A Unique Late Triassic Dinosauromorph Assemblage Reveals Dinosaur Ancestral Anatomy and Diet". Current Biology. 26 (22): 3090–3095. doi:10.1016/j.cub.2016.09.040. PMID 27839975.
- ^ Müller, Rodrigo Temp; Garcia, Maurício Silva (August 26, 2020). "A paraphyletic 'Silesauridae' as an alternative hypothesis for the initial radiation of ornithischian dinosaurs". Biology Letters. 16 (8): 20200417. doi:10.1098/rsbl.2020.0417. PMC 7480155. PMID 32842895.
- ^ Alfaro, M.E., F. Santini, C. Brock, H. Alamillo, A. Dornburg. D.L. Rabosky, G. Carnevale, and L.J. Harmon (2009). "Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates". Proceedings of the National Academy of Sciences USA. 106 (32): 13410–13414. doi:10.1073/pnas.0811087106. PMC 2715324. PMID 19633192.
{{cite journal}}: CS1 maint: PMC format (link) CS1 maint: multiple names: authors list (link) - ^ Wang, S.C., and Dodson, P. (2006). "Estimating the Diversity of Dinosaurs". Proceedings of the National Academy of Sciences USA. 103 (37): 13601–13605. doi:10.1073/pnas.0606028103. PMC 1564218. PMID 16954187.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Will the real dinosaurs stand up?, BBC, September 17, 2008
- ^ أ ب ت مكليود, إن, روسون, بي إف, فوري, بي إل, بانر, إف تي, بوداغر-فاضل, إم كيه, بون, بي آر, برنيت, جيه إيه, تشيمبرز, بي, كلفر, إس, إيفانز, إس إي, جيفري, سي, كامنسكي, إم إيه, لورد, إيه آر, ميلنر, إيه سي, ميلنر, إيه آر, موريس, إن, أوين, إي, روزن, بي آر, سميث, إيه بي, تايلور, PD, أوركوهارت, إي ويونغ, جيه آر (1997). "التحول الأحيائي الثلاثي-طباشيري". جورنال أوف جيولوجيكال سوسايتي. 154 (2): 265–292. doi:10.1144/gsjgs.154.2.0265.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "MacLeod" معرف أكثر من مرة بمحتويات مختلفة. - ^ باكر, ر. ت., غالتون, ب., 1974. أحادية أرومة الديناصورات وعصر الفقاريات الجديد. نيتشر 248:168-172.
- ^ أ ب ت ث بنتون, مايكل جيه. (2004). "أصل الديناصورات وعلاقاتها". In وايشامبل, ديفد بي.; دودسون, بيتر; وأوسملسكا, هالزكا (المحررون.) (ed.). الديناصورات (الطبعة الثانية ed.). بيركلي: مطبعة جامعة كاليفورنيا. pp. 7–19. ISBN 0-520-24209-2.
{{cite book}}: CS1 maint: multiple names: editors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "MJB04dino" معرف أكثر من مرة بمحتويات مختلفة. - ^ أولشفسكي, جي. (2000).
"قائمة تحقق مشروحة لأنواع الديناصورات حسب القارات." التضاريس الوسطى, 3: 1–157 خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "olshevsky2000" معرف أكثر من مرة بمحتويات مختلفة. - ^ أ ب ت ث ج ح خ باديان, كيه. (2004). باسال أفيالاي. المحدودة: وايشامبل, دي.بي., دودسون, بي., وأوسملسكا, إتش. (المحررون.). الديناصورات (الطبعة الثانية). مطبعة جامعة كاليفورنيا:بيركلي, 210–231. ISBN 0-520-24209-2. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "KP04" معرف أكثر من مرة بمحتويات مختلفة. - ^ غلت, دونالد إف. (1997). الديناصورات: الموسوعة. جيفرسون, نورث كارولينا: مكفارلاند وشركاه. p. 40. ISBN 0-89950-917-7. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "DFG97" معرف أكثر من مرة بمحتويات مختلفة. - ^ لامبرت, ديفد (1990). كتاب بيانات الديناصورات. نيويورك: آفون بوكس. p. 288. ISBN 0-380-75896-3.
{{cite book}}: Check|isbn=value: checksum (help); Unknown parameter|coauthors=ignored (|author=suggested) (help) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "DL90" معرف أكثر من مرة بمحتويات مختلفة. - ^ موراليس, مايكل (1997). "الفقاريات غير الديناصورية في الحقبة الوسطى". In فارلو, جيمس أو.; وبريت-سورمان, مايكل كيه. (المحررون.) (ed.). The Complete Dinosaur. بلومنغتون: مطبعة جامعة إنديانا. pp. 607–624. ISBN 0-253-33349-0.
{{cite book}}: CS1 maint: multiple names: editors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "MM97" معرف أكثر من مرة بمحتويات مختلفة. - ^ هو ياومنغ (2005). "ثدييات حقبة وسطى كبيرة تتغذى على الديناصورات". نيتشر. 433: 149–152. doi:10.1038/nature03102.
{{cite journal}}: Unknown parameter|coauthors=ignored (|author=suggested) (help) - ^ وانغ, إس. سي., and دودسون, بي. (2006). "تقدير تنوع الديناصورات". بروسيدينغز أوف ذا ناشنال أكاديمي. 103 (37): 13601–13605. doi:10.1073/pnas.0606028103. PMID 16954187.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ رسل, ديل إيه. (1995). "الصين وعالم حقبة الديناصورات الضائع". هيستوريكال بيولوجي. 10: 3–12. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "russell1995" معرف أكثر من مرة بمحتويات مختلفة. - ^ هل ستصمد الديناصورات الحقيقية؟, BBC, September 17, 2008
- ^ بنتون, مايكل جيه. (2004). "أصول الديناصورات وعلاقاتها". In وايشامبل, ديفد بي.; دودسون, بيتر; وأوسملسكا, هالزكا(المحررون.) (ed.). الديناصورات (2nd ed.). بيركلي: مطبعة جامعة كاليفورنيا. pp. 7–19. ISBN 0-520-24209-2.
{{cite book}}: CS1 maint: multiple names: editors list (link) - ^ Seeley, H.G. (1887). "On the classification of the fossil animals commonly named Dinosauria". Proc R Soc London. Royal Society. 43: 165–171. doi:10.1098/rspl.1887.0117.
- ^ Romer, A.S. (1956). Osteology of the Reptiles. University of Chicago. ISBN 089464985X.
- ^ Ostrom, J.H. (1980). "The evidence of endothermy in dinosaurs". In Thomas, R.D.K. and Olson, E.C. (ed.). A cold look at the warm-blooded dinosaurs. Boulder, CO: American Association for the Advancement of Science. pp. 82–105.
{{cite book}}: CS1 maint: multiple names: editors list (link) - ^ Bakker, R. T., and Galton, P. (1974). "Dinosaur monophyly and a new class of vertebrates". Nature. 248: 168–172. doi:10.1038/248168a0.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Kump LR, Pavlov A & Arthur MA (2005). "Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia". Geology. 33 (5): 397–400. doi:10.1130/G21295.1.
- ^ Tanner LH, Lucas SG & Chapman MG (2004). "Assessing the record and causes of Late Triassic extinctions" (PDF). Earth-Science Reviews. 65 (1–2): 103–139. doi:10.1016/S0012-8252(03)00082-5. Archived from the original (PDF) on October 25, 2007. Retrieved 2007-10-22. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "TannerLucas" معرف أكثر من مرة بمحتويات مختلفة. - ^ أ ب Sereno PC (1999). "The evolution of dinosaurs". Science. 284 (5423): 2137–2147. doi:10.1126/science.284.5423.2137. PMID 10381873.
- ^ Sereno, P.C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (1993). "Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria". Nature. 361: 64–66. doi:10.1038/361064a0. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "SFRM93" معرف أكثر من مرة بمحتويات مختلفة. - ^ Brusatte, S.L.; Benton, MJ; Ruta, M; Lloyd, GT (2008). "Superiority, competition, and opportunism in the evolutionary radiation of dinosaurs". Science. 321 (5895): 1485–1488. doi:10.1126/science.1161833. PMID 18787166.
- ^ أ ب Lloyd, G.T., Davis, K.E., Pisani, D (22 July 2008). "Dinosaurs and the Cretaceous Terrestrial Revolution". Proceedings of the Royal Society: Biology. 275 (1650): 2483–90. doi:10.1098/rspb.2008.0715. PMC 2603200. PMID 18647715. Retrieved 2008-07-28.
{{cite journal}}: CS1 maint: multiple names: authors list (link) The URL contains links to all the content, and it's all free. - ^ أ ب ت ث Holtz, Thomas R., Jr. (2004). "Mesozoic biogeography of Dinosauria". In Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.) (ed.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 627–642. ISBN 0-520-24209-2.
{{cite book}}:|editor=has generic name (help); Unknown parameter|coauthors=ignored (|author=suggested) (help)CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "HCL04" معرف أكثر من مرة بمحتويات مختلفة. - ^ أ ب ت ث ج Fastovsky, David E. (2004). "Dinosaur paleoecology". In Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.) (ed.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 614–626. ISBN 0-520-24209-2.
{{cite book}}:|editor=has generic name (help); Unknown parameter|coauthors=ignored (|author=suggested) (help)CS1 maint: multiple names: editors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "FS04" معرف أكثر من مرة بمحتويات مختلفة. - ^ Sereno, P.C.; Wilson, JA; Witmer, LM; Whitlock, JA; Maga, A; Ide, O; Rowe, TA; Kemp, Tom (2007). "Structural extremes in a Cretaceous dinosaur". PLoS ONE. 2 (11): e1230. doi:10.1371/journal.pone.0001230. PMC 2077925. PMID 18030355.
{{cite journal}}: CS1 maint: unflagged free DOI (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "serenoetal07" معرف أكثر من مرة بمحتويات مختلفة. - ^ Prasad, V.; Strömberg, CA; Alimohammadian, H; Sahni, A (2005). "Dinosaur coprolites and the early evolution of grasses and grazers". Science. 310 (5751): 1170–1180. doi:10.1126/science.1118806. PMID 16293759. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "PSAS05" معرف أكثر من مرة بمحتويات مختلفة. - ^ Archibald, J. David (2004). "Dinosaur Extinction". In Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.) (ed.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 672–684. ISBN 0-520-24209-2.
{{cite book}}:|editor=has generic name (help); Unknown parameter|coauthors=ignored (|author=suggested) (help)CS1 maint: multiple names: editors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "AF04" معرف أكثر من مرة بمحتويات مختلفة. - ^ Benton, M.J. (2004). Vertebrate Paleontology. Blackwell Publishers. xii–452. ISBN 0-632-05614-2.
{{cite book}}: Unknown parameter|nopp=ignored (|no-pp=suggested) (help) - ^ Paul, G.S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster. pp. 248–250. ISBN 0671619462. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "GSP88" معرف أكثر من مرة بمحتويات مختلفة. - ^ Clark, J.M., Maryanska, T., and Barsbold, R. (2004). "Therizinosauroidea", in The Dinosauria, 2nd ed. 151–164. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "clarketal2004" معرف أكثر من مرة بمحتويات مختلفة. - ^ Norell, M.A., and Makovicky, P.J. (2004). "Dromaeosauridae", in The Dinosauria, 2nd ed. 196–210. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "MAPM04" معرف أكثر من مرة بمحتويات مختلفة. - ^ الديناصورات، الموسوعة المعرفية الشاملة
- ^ أ ب Dal Sasso, C. and Signore, M. (1998)). "Exceptional soft-tissue preservation in a theropod dinosaur from Italy". Nature. 292 (6674): 383–387. doi:10.1038/32884.
{{cite journal}}: Check date values in:|date=(help)CS1 maint: multiple names: authors list (link) - ^ أ ب ت Schweitzer, M.H., Wittmeyer, J.L. and Horner, J.R. (2005). "Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex". Science. 307 (5717): 1952–1955. doi:10.1126/science.1108397. PMID 15790853.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "Schweitzer2005" معرف أكثر من مرة بمحتويات مختلفة. - ^ أ ب ت استكشاف بيئة الحقبة الوسطى. تاريخ الولوج 13-03-2011.
- ^ أ ب ت ث ج ح خ د ذ ر كتب غوغل "The Evolution And Extinction of Dinosaurs" (تطور وانقراض الديناصورات). لـ"David B. Weishampel" و"David E. Fastovsky". ص387-402. تاريخ الولوج 13-03-2011.
- ^ أ ب صور الأخبار: اكتشاف ديناصور عملاق في أنتاركتيكا. ناشيونال جيوغرافيك. تاريخ الولوج 13-03-2011.
- ^ برفقة الديناصورات: الديناصورات الأسترالية. تاريخ الولوج 13-03-2011.
- ^ الديناصورات الأسترالية. تاريخ الولوج 13-03-2011.
- ^ انتشار الديناصورات الجغرافي. تاريخ الولوج 13-03-2011.
- ^ زواحف وديناصورات العصر الجوراسي. تاريخ الولوج 13-03-2011.
- ^ أ ب ت ث كتاب "the Ultimate Dinosaur Book" لـ"David Lambert"، ص14-19، من دار "Dorling Kindersley (DK)" في لندن.
- ^ العصر الطباشيري:انقراض الديناصورات. تاريخ الولوج 19 مارس 2010.
- ^ أ ب ما هي المواطن التي عاشت فيها الديناصورات؟. تاريخ الولوج 15-03-2011.
- ^ أ ب ت التاريخ الطبيعي: الديناصورات (زواحف منقرضة) - الموطن. الموسوعة البريطانية. تاريخ الولوج 15-03-2011.
- ^ احتوت بيئات الديناصورات أنوعاً أكثر مما اعتقد سابقاً. تاريخ الولوج 15-03-2011.
- ^ ديناصورات صحراء الولايات المتحدة. تاريخ الولوج 15-03-2011.
- ^ أ ب ت مواطن الديناصورات الطبيعية. تاريخ الولوج 15-03-2011.
- ^ أ ب ت برفقة الديناصور - الخط الزمني: قبل 220 مليون عام. من هيئة الإذاعة البريطانية. تاريخ الولوج 15-03-2011.
- ^ أ ب كتاب "قاموس الديناصورات: إنكليزي - فرنسي - عربي"، من "دار المجاني".
- ^ أ ب برفقة الديناصورات: خط زمني: قبل 106 مليون سنة. بي بي سي. تاريخ الولوج 15-03-2011.
- ^ موطن التي ريكس الطبيعي - موقع اسأل بيولوجياً. تاريخ الولوج 15-03-2011.
- ^ "Assessment and Update Status Report on the Blue Whale Balaenoptera musculus" (PDF). Committee on the Status of Endangered Wildlife in Canada. 2002. Archived from the original (PDF) on December 2, 2007. Retrieved 2007-12-05.
- ^ أ ب ت Carpenter, Kenneth (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus". In Foster, John R.; and Lucas, Spencer G. (eds.) (ed.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque: New Mexico Museum of Natural History and Science. pp. 131–138.
{{cite book}}:|editor=has generic name (help)CS1 maint: multiple names: editors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "KC06" معرف أكثر من مرة بمحتويات مختلفة. - ^ Farlow, James A. (1993). "On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large, carnivorous dinosaurs". Functional Morphology and Evolution. American Journal of Science, Special Volume 293-A. pp. 167–199.
{{cite book}}: Unknown parameter|editors=ignored (|editor=suggested) (help) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "JF93" معرف أكثر من مرة بمحتويات مختلفة. - ^ Peczkis, J. (1994). "Implications of body-mass estimates for dinosaurs". Journal of Vertebrate Paleontology. 14 (4): 520–33. doi:10.1080/02724634.1995.10011575.
- ^ "Anatomy and evolution". National Museum of Natural History. Retrieved 2007-11-21.
- ^ Colbert, E.H. (1968). Men and Dinosaurs: The Search in Field and Laboratory. E. P. Dutton & Company:New York, vii + 283 p. ISBN 0-14-021288-4. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "EC68" معرف أكثر من مرة بمحتويات مختلفة. - ^ Lovelace, David M. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional. 65 (4): 527–544. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "LHW07" معرف أكثر من مرة بمحتويات مختلفة. - ^ dal Sasso, C., Maganuco, S., Buffetaut, E., and Mendez, M.A. (2006). New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities. Journal of Vertebrate Paleontology 25(4):888–896. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "SMBM06" معرف أكثر من مرة بمحتويات مختلفة. - ^ Therrien, F. (2007). "My theropod is bigger than yours...or not: estimating body size from skull length in theropods". Journal of Vertebrate Paleontology. 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
{{cite journal}}: Unknown parameter|coauthors=ignored (|author=suggested) (help) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "TH07" معرف أكثر من مرة بمحتويات مختلفة. - ^ أ ب Zhang, Fucheng; Zhou, Zhonghe; Xu, Xing; Wang, Xiaolin and Sullivan, Corwin. "A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers" Nature 455, 1105–1108 (23 October 2008) | doi:10.1038/nature07447
- ^ أ ب Xu, X., Zhao, Q., Norell, M., Sullivan, C., Hone, D., Erickson, G., Wang, X., Han, F. and Guo, Y. (2009). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin." Chinese Science Bulletin, six pages, accepted November 15, 2008. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "anchiadvance" معرف أكثر من مرة بمحتويات مختلفة. - ^ Holtz, Thomas R. Jr. (2008) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages Supplementary Information
خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "Holtz2008" معرف أكثر من مرة بمحتويات مختلفة. - ^ Butler, R.J.; Zhao, Q. (2009). "The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China". Cretaceous Research. 30 (1): 63–77. doi:10.1016/j.cretres.2008.03.002. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "butler&zhao2009" معرف أكثر من مرة بمحتويات مختلفة. - ^ Yans J, Dejax J, Pons D, Dupuis C & Taquet P (2005). "Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)". Comptes Rendus Palevol (in French). 4 (1–2): 135–150. doi:10.1016/j.crpv.2004.12.003.
{{cite journal}}: CS1 maint: multiple names: authors list (link) CS1 maint: unrecognized language (link) - ^ Day, J.J.; Upchurch, P; Norman, DB; Gale, AS; Powell, HP (2002). "Sauropod trackways, evolution, and behavior". Science. 296 (5573): 1659. doi:10.1126/science.1070167. PMID 12040187.
- ^ Wright, Joanna L. (2005). "Steps in understanding sauropod biology". In Curry Rogers, Kristina A.; and Wilson, Jeffrey A. (ed.). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. pp. 252–284. ISBN 0-520-24623-3.
{{cite book}}: CS1 maint: multiple names: editors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "JLW05" معرف أكثر من مرة بمحتويات مختلفة. - ^ Varricchio, D.J.; Sereno, Paul C.; Xijin, Zhao; Lin, Tan; Wilson, Jeffery A.; Lyon, Gabrielle H. (2008). "Mud-trapped herd captures evidence of distinctive dinosaur sociality" (PDF). Acta Palaeontologica Polonica. 53 (4): 567–578. doi:10.4202/app.2008.0402. Retrieved 2008-12-09. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "DVetal08sino" معرف أكثر من مرة بمحتويات مختلفة. - ^ Lessem, Don (1993). "Allosaurus". The Dinosaur Society's Dinosaur Encyclopedia. Random House. pp. 19–20. ISBN 0-679-41770-2.
{{cite book}}: Unknown parameter|coauthors=ignored (|author=suggested) (help) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "LG93" معرف أكثر من مرة بمحتويات مختلفة. - ^ Maxwell, W. D.; Ostrom, John (1995). "Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations". Journal of Vertebrate Paleontology. 15 (4): 707–712. doi:10.1080/02724634.1995.10011256.(abstract) خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "maxwell&ostrom1995" معرف أكثر من مرة بمحتويات مختلفة. - ^ Roach, Brian T.; Brinkman, Daniel L. (2007). "A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs". Bulletin of the Peabody Museum of Natural History. 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ^ Horner, J.R.; Makela, Robert (1979). "Nest of juveniles provides evidence of family structure among dinosaurs". Nature. 282 (5736): 296–298. doi:10.1038/282296a0. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "HM79" معرف أكثر من مرة بمحتويات مختلفة. - ^ Chiappe, Luis M. (2005). "Nesting titanosaurs from Auca Mahuevo and adjacent sites". In Curry Rogers, Kristina A.; and Wilson, Jeffrey A. (ed.). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. pp. 285–302. ISBN 0-520-24623-3.
{{cite book}}: Unknown parameter|coauthors=ignored (|author=suggested) (help)CS1 maint: multiple names: editors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "LMCetal05" معرف أكثر من مرة بمحتويات مختلفة. - ^ Oviraptor nesting Oviraptor nests or Protoceratops?
- ^ Meng Qingjin; Liu Jinyuan; Varricchio, David J.; Huang, Timothy; and Gao Chunling (2004). "Parental care in an ornithischian dinosaur". Nature. 431 (7005): 145–146. doi:10.1038/431145a. PMID 15356619.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Reisz RR, Scott, D Sues, H-D, Evans, DC & Raath, MA (2005). "Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance". Science. 309 (5735): 761–764. doi:10.1126/science.1114942. PMID 16051793.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "Reiszetal05" معرف أكثر من مرة بمحتويات مختلفة. - ^ Dinosaur family tracks Footprints show maternal instinct after leaving the nest.
- ^ Tanke, Darren H. (1998). "Head-biting behavior in theropod dinosaurs: paleopathological evidence" (PDF). Gaia (15): 167–184. ISSN 0871-5424. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "PC98" معرف أكثر من مرة بمحتويات مختلفة. - ^ "The Fighting Dinosaurs". American Museum of Natural History. Retrieved 2007-12-05.
- ^ أ ب Carpenter, K. (1998). "Evidence of predatory behavior by theropod dinosaurs". Gaia. 15: 135–144. Retrieved 2007-12-05. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "carpenter1998" معرف أكثر من مرة بمحتويات مختلفة. - ^ Rogers, Raymond R.; Krause, DW; Curry Rogers, K (2007). "Cannibalism in the Madagascan dinosaur Majungatholus atopus". Nature. 422 (6931): 515–518. doi:10.1038/nature01532. PMID 12673249.
- ^ Varricchio DJ, Martin, AJ and Katsura, Y (2007). "First trace and body fossil evidence of a burrowing, denning dinosaur". Proceedings of the Royal Society B: Biological Sciences. 274 (1616): 1361–1368. doi:10.1098/rspb.2006.0443. PMC 2176205. PMID 17374596.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "VMK07" معرف أكثر من مرة بمحتويات مختلفة. - ^ Chatterjee, S.; Templin, R. J. (2007). "Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui" (PDF). Proceedings of the National Academy of Sciences. 104 (5): 1576–1580. doi:10.1073/pnas.0609975104. PMC 1780066. PMID 17242354. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "chatterjee2007" معرف أكثر من مرة بمحتويات مختلفة. - ^ Alexander RM (2006). "Dinosaur biomechanics". Proceedings of the Royal Society of Biological Sciences. 273 (1596): 1849–1855. doi:10.1098/rspb.2006.3532. PMC 1634776. PMID 16822743.
- ^ Goriely A & McMillen T (2002). "Shape of a cracking whip". Physical Review Letters. 88 (24): 244301. doi:10.1103/PhysRevLett.88.244301. PMID 12059302.
- ^ Henderson, D.M. (2003). "Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: A computational analysis". Canadian Journal of Zoology. 81 (8): 1346–1357. doi:10.1139/z03-122.
- ^ أ ب ت ث ج Senter, P. (2008). "Voices of the past: a review of Paleozoic and Mesozoic animal sounds". Historical Biology. 20 (4): 255–287. doi:10.1080/08912960903033327.
- ^ Hopson, James A. (1975). "The evolution of cranial display structures in hadrosaurian dinosaurs". Paleobiology. 1 (1): 21–43. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "JAH75" معرف أكثر من مرة بمحتويات مختلفة. - ^ Diegert, Carl F. (1998). "A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen". Journal of Vertebrate Paleontology. 18 (3, Suppl.): 38A. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "DW98" معرف أكثر من مرة بمحتويات مختلفة. - ^ Parsons, K.M. (2001). Drawing Out Leviathan. Indiana University Press. 22–48. ISBN 0-253-33937-5.
- ^ Fisher, P. E., Russell, D. A., Stoskopf, M. K., Barrick, R. E., Hammer, M. & Kuzmitz, A. A. (2000). "Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur". Science. 288 (5465): 503–505. doi:10.1126/science.288.5465.503. PMID 10775107.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Hillenius, W. J. & Ruben, J. A. (2004). "The evolution of endothermy in terrestrial vertebrates: Who? when? why?". Physiological and Biochemical Zoology. 77 (6): 1019–1042. doi:10.1086/425185. PMID 15674773.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Rowe T, McBride EF, & Sereno PC (2001). "Dinosaur with a Heart of Stone". Science. 291 (5505): 783. doi:10.1126/science.291.5505.783a. PMID 11157158.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Chinsamy, Anusuya; and Hillenius, Willem J. (2004). "Physiology of nonavian dinosaurs". The Dinosauria, 2nd. 643–659. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "CH04" معرف أكثر من مرة بمحتويات مختلفة. - ^ Huxley, Thomas H. (1868). "On the animals which are most nearly intermediate between birds and reptiles". Annals of the Magazine of Natural History. 4 (2): 66–75.
- ^ Heilmann, Gerhard (1926). The Origin of Birds. London: Witherby. pp. 208pp. ISBN 0-486-22784-7. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "heilmann" معرف أكثر من مرة بمحتويات مختلفة. - ^ Osborn, Henry Fairfield (1924). "Three new Theropoda, Protoceratops zone, central Mongolia" (PDF). American Museum Novitates. 144: 1–12.
- ^ Ostrom, John H. (1973). "The ancestry of birds". Nature. 242 (5393): 136. doi:10.1038/242136a0. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "ostrom1973" معرف أكثر من مرة بمحتويات مختلفة. - ^ Gauthier, Jacques. (1986). "Saurischian monophyly and the origin of birds". In Padian, Kevin. (ed.) (ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8. pp. 1–55.
{{cite book}}:|editor=has generic name (help) - ^ Holtz, Jr., T.R. (2000). "Classification and evolution of the dinosaur groups". In Paul, G.S. (ed.). The Scientific American Book of Dinosaurs. St. Martin's Press. pp. 140–168. ISBN 0-312-26226-4.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ^ Mayr, G., Pohl, B. and Peters, D.S. (2005). "A Well-Preserved Archaeopteryx Specimen with Theropod Features". Science. 310 (5753): 1483–1486. doi:10.1126/science.1120331. PMID 16322455.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Martin, Larry D. (2006). "A basal archosaurian origin for birds". Acta Zoologica Sinica. 50 (6): 977–990.
- ^ أ ب Feduccia, A. (2002). "Birds are dinosaurs: simple answer to a complex problem". The Auk. 119: 1187–1201. doi:10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "AF02" معرف أكثر من مرة بمحتويات مختلفة. - ^ Wellnhofer, P (1988). "Ein neuer Exemplar von Archaeopteryx". Archaeopteryx. 6: 1–30.
- ^ Xu X.; Norell, M.A.; Kuang X.; Wang X.; Zhao Q.; and Jia C. (2004). "Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids". Nature. 431 (7009): 680–684. doi:10.1038/nature02855. PMID 15470426.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Göhlich, U.B.; Chiappe, LM (2006). "A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago". Nature. 440 (7082): 329–332. doi:10.1038/nature04579. PMID 16541071. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "GC06" معرف أكثر من مرة بمحتويات مختلفة. - ^ Lingham-Soliar, T. (2003). "The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers". Naturwissenschaften. 12 (12): 563–567. doi:10.1007/s00114-003-0483-7. PMID 14676953. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "TLS03" معرف أكثر من مرة بمحتويات مختلفة. - ^ أ ب Feduccia, A.; Lingham-Soliar, T; Hinchliffe, JR (2005). "Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence". Journal of Morphology. 266 (2): 125–166. doi:10.1002/jmor.10382. PMID 16217748.
- ^ Lingham-Soliar, T.; Feduccia, A; Wang, X (2007). "A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres". Proceedings of the Biological Sciences. 274 (1620): 1823–9. doi:10.1098/rspb.2007.0352. PMC 2270928. PMID 17521978. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "LSFX07" معرف أكثر من مرة بمحتويات مختلفة. - ^ Prum, Richard O. (2003). <550:ACCOTT>2.0.CO;2-0 "Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002". The Auk. 120 (2): 550–61. doi:10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2.
{{cite journal}}: Unknown parameter|month=ignored (help) - ^ O'Connor, P.M. & Claessens, L.P.A.M. (2005). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature. 436 (7048): 253–256. doi:10.1038/nature03716. PMID 16015329.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Sereno, P.C.; Martinez, RN; Wilson, JA; Varricchio, DJ; Alcober, OA; Larsson, HC; Kemp, Tom (2008). "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina". PLoS ONE. 3 (9): e3303. doi:10.1371/journal.pone.0003303. PMC 2553519. PMID 18825273. Retrieved 2008-10-27.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: unflagged free DOI (link) - ^ Meat-Eating Dinosaur from Argentina Had Bird-Like Breathing System Newswise, Retrieved on September 29, 2008.
- ^ Wings O (2007). "A review of gastrolith function with implications for fossil vertebrates and a revised classification" (PDF). Palaeontologica Polonica. 52 (1): 1–16.
- ^ Xu, X. and Norell, M.A. (2004). "A new troodontid dinosaur from China with avian-like sleeping posture". Nature. 431 (7010): 838–841. doi:10.1038/nature02898. PMID 15483610.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Miller KG, Kominz MA, Browning JV, Wright JD, Mountain GS, Katz ME, Sugarman PJ, Cramer BS, Christie-Blick N, Pekar SF (2005). "The Phanerozoic record of global sea-level change". Science. 310 (5752): 1293–8. doi:10.1126/science.1116412. PMID 16311326.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "pmid16311326" معرف أكثر من مرة بمحتويات مختلفة. - ^ McArthura JM, Janssenb NMM, Rebouletc S, Lengd MJ, Thirlwalle MF & van de Shootbruggef B (2007). "Palaeotemperatures, polar ice-volume, and isotope stratigraphy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): The Early Cretaceous (Berriasian, Valanginian, Hauterivian)". Palaeogeography, Palaeoclimatology, Palaeoecology. 248 (3–4): 391–430. doi:10.1016/j.palaeo.2006.12.015.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Alvarez, LW, Alvarez, W, Asaro, F, and Michel, HV (1980). "Extraterrestrial cause for the Cretaceous–Tertiary extinction". Science. 208 (4448): 1095–1108. doi:10.1126/science.208.4448.1095. PMID 17783054.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "Alvarezetal80" معرف أكثر من مرة بمحتويات مختلفة. - ^ Hildebrand, Alan R.; Penfield, Glen T.; Kring, David A.; Pilkington, Mark; Zanoguera, Antonio Camargo; Jacobsen, Stein B.; Boynton, William V. (1991). "Chicxulub Crater; a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico". Geology. 19 (9): 867–871. doi:10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "ARHetal91" معرف أكثر من مرة بمحتويات مختلفة. - ^ Pope KO, Ocampo AC, Kinsland GL, Smith R (1996). "Surface expression of the Chicxulub crater". Geology. 24 (6): 527–30. doi:10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2. PMID 11539331.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "KPetal96" معرف أكثر من مرة بمحتويات مختلفة. - ^ P, Claeys; Goderis, S (2007-09-05). "Solar System: Lethal billiards". Nature. 449 (7158): 30–31. doi:10.1038/449030a. PMID 17805281.
- ^ أ ب edited by Christian Koeberl and Kenneth G. MacLeod. (2002). Catastrophic Events and Mass Extinctions. Geological Society of America. ISBN 0-8137-2356-6. OCLC 213836505.
{{cite book}}:|author=has generic name (help); Unknown parameter|unused_data=ignored (help) - ^ أ ب Hofman, C, Féraud, G & Courtillot, V (2000). "40Ar/39Ar dating of mineral separates and whole rocks from the Western Ghats lava pile: further constraints on duration and age of the Deccan traps". Earth and Planetary Science Letters. 180: 13–27. doi:10.1016/S0012-821X(00)00159-X.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "wwnjsd" معرف أكثر من مرة بمحتويات مختلفة. - ^ أ ب ت Duncan, RA & Pyle, DG (1988). "Rapid eruption of the Deccan flood basalts at the Cretaceous/Tertiary boundary". Nature. 333: 841–843. doi:10.1038/333841a0.
{{cite journal}}: CS1 maint: multiple names: authors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "Duncan" معرف أكثر من مرة بمحتويات مختلفة. - ^ Alvarez, W (1997). T. rex and the Crater of Doom. Princeton University Press. pp. 130–146. ISBN 978-0-691-01630-6.
- ^ خطأ استشهاد: وسم
<ref>غير صحيح؛ لا نص تم توفيره للمراجع المسماةfassett2011 - ^ خطأ استشهاد: وسم
<ref>غير صحيح؛ لا نص تم توفيره للمراجع المسماةfassett2009 - ^ خطأ استشهاد: وسم
<ref>غير صحيح؛ لا نص تم توفيره للمراجع المسماةsloan1986 - ^ Lucas, S.G.; Sullivan, R.M.; Cather, S.M.; Jasinski, S.E.; Fowler, D.W.; Heckert, A.B.; Spielmann, J.A.; Hunt, A.P. (2009). "No definitive evidence of Paleocene dinosaurs in the San Juan Basin". Palaeontologia Electronica. 12 (2): 8A.
- ^ Renne, P.R.; Goodwin, M.B. (2012). "Direct U-Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico: COMMENT". Geology. 40 (4): e259. Bibcode:2012Geo....40E.259R. doi:10.1130/G32521C.1.
- ^ Lofgren, D.L.; Hotton, C.L.; Runkel, A.C. (1990). "Reworking of Cretaceous dinosaurs into Paleocene channel, deposits, upper Hell Creek Formation, Montana". Geology. 18 (9): 874–877. Bibcode:1990Geo....18..874L. doi:10.1130/0091-7613(1990)018<0874:ROCDIP>2.3.CO;2.
- ^ Koenig, A.E.; Lucas, S.G.; Neymark, L.A.; Heckert, A.B.; Sullivan, R.M.; Jasinski, S.E.; Fowler, D.W. (2012). "Direct U-Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico: COMMENT". Geology. 40 (4): e262. Bibcode:2012Geo....40E.262K. doi:10.1130/G32154C.1.
- ^ Kaye, T. G.; Gaugler, G; Sawlowicz, Z; Stepanova, Anna (2008). "Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms". PLoS ONE. 3 (7): e2808. doi:10.1371/journal.pone.0002808. PMC 2483347. PMID 18665236. Retrieved 2008-10-27.
{{cite journal}}: Unknown parameter|month=ignored (help)CS1 maint: unflagged free DOI (link) - ^ New Research Challenges Notion That Dinosaur Soft Tissues Still Survive Newswise, Retrieved on 29 July 2008.
- ^ Wang, H., Yan, Z. and Jin, D. (1 May 1997). "Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil". Molecular Biology and Evolution. 14 (5): 589–591. PMID 9159936. Retrieved 2007-12-05.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Chang BS, Jönsson K, Kazmi MA, Donoghue MJ, Sakmar TP (1 September 2002). "Recreating a Functional Ancestral Archosaur Visual Pigment". Molecular Biology and Evolution. 19 (9): 1483–1489. PMID 12200476. Retrieved 2007-12-05.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Schweitzer MH, Marshall M, Carron K, Bohle DS, Busse SC, Arnold EV, Barnard D, Horner JR, Starkey JR (1997). "Heme compounds in dinosaur trabecular bone". Proc Natl Acad Sci U S A. 94 (12): 6291–6. doi:10.1073/pnas.94.12.6291. PMC 21042. PMID 9177210.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Embery G, Milner AC, Waddington RJ, Hall RC, Langley MS, Milan AM (2003). "Identification of proteinaceous material in the bone of the dinosaur Iguanodon". Connect Tissue Res. 44 (Suppl 1): 41–6. doi:10.1080/713713598. PMID 12952172.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ "حقائق مثيرة داخل بيضة ديناصور عمرها 70 مليون سنة".
- ^ "Definition of dinosaur" Merriam-Webster's Online Dictionary. Accessed 26 May 2007. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "m-w" معرف أكثر من مرة بمحتويات مختلفة. - ^ Torrens, H.S. (1993). "The dinosaurs and dinomania over 150 years". Modern Geology. 18 (2): 257–286.
- ^ Breithaupt, Brent H. (1997). "First golden period in the USA". In Currie, Philip J. and Padian, Kevin (eds.) (ed.). The Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 347–350. ISBN 978-0-12-226810-6.
{{cite book}}:|editor=has generic name (help)CS1 maint: multiple names: editors list (link) خطأ استشهاد: وسم<ref>غير صالح؛ الاسم "breithaupt1997" معرف أكثر من مرة بمحتويات مختلفة. - ^ "London. Michaelmas term lately over, and the Lord Chancellor sitting in Lincoln's Inn Hall. Implacable November weather. As much mud in the streets, as if the waters had but newly retired from the face of the earth, and it would not be wonderful to meet a Megalosaurus, forty feet long or so, waddling like an elephantine lizard up Holborne Hill." From page 1 of Dickens, Charles J.H. (1852). Bleak House. London: Bradbury & Evans. خطأ استشهاد: وسم
<ref>غير صالح؛ الاسم "bleakhouse" معرف أكثر من مرة بمحتويات مختلفة. - ^ Glut, D.F., and Brett-Surman, M.K. (1997). Farlow, James O. and Brett-Surman, Michael K. (eds.) (ed.). The Complete Dinosaur. Indiana University Press. pp. 675–697. ISBN 978-0-253-21313-6.
{{cite book}}:|editor=has generic name (help)CS1 maint: multiple names: authors list (link)
خطأ استشهاد: الوسم <ref> ذو الاسم "alexander2006" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Alfaroetal2009" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Alvarez1997" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "AMNH" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "AmosBBC" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "BBCdinobonemed" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "BBCtracks" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "B012" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "buckland1824" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "changetal2002" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Day2002" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "dong1992" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "DS02" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Emberyatal2003" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Fassett" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "fassettetall2011" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "FBS97" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "FS05" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "GM25" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "goriely" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "G45" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "HDS97" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Henderson2006" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Holland1909" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Holmes" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "KPA" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "LARB99" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Letal05" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "L99" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "LSJ" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "LW08" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Mayretal2005" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "McAetal2007" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "nesbitt2011" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "newswise2" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "NIP07" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "OConnorClaessens2005" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Owen1842" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "parsons2001" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Peczkis1994" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Prum2003" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "RC05" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "RMetal10" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "SchmitzMotani2011" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Schweitzeretal1997" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "search.eb" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "senter2008" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Sereno2008" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Sloan" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Sullivan" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "TKMB07" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Wang&Dodson" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Wangetal1997" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "WAS97" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "wings2007" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Xuetal2004" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "XUNorell2004" المُعرّف في <references> غير مستخدم في النص السابق.
خطأ استشهاد: الوسم <ref> ذو الاسم "Yans" المُعرّف في <references> غير مستخدم في النص السابق.
<ref> ذو الاسم "zhang2008" المُعرّف في <references> غير مستخدم في النص السابق.قراءات إضافية
| مراجع مكتبية عن Dinosaurs |
- Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction. New York: Morrow. ISBN 0-688-04287-2.
- Holtz, Thomas R. Jr. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. New York: Random House. ISBN 978-0-375-82419-7.
- Paul, Gregory S. (2000). The Scientific American Book of Dinosaurs. New York: St. Martin's Press. ISBN 0-312-26226-4.
- Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: The Johns Hopkins University Press. ISBN 0-8018-6763-0.
- Sternberg, C. M. (1966). Canadian Dinosaurs, in Geological Series, no. 54. Second ed. [Ottawa]: National Museum of Canada. 28 p., amply ill.
- Stewart, Tabori & Chang (1997). "The Humongous Book of Dinosaurs". New York: Stewart, Tabori & Chang. Retrieved May 17, 2012. ISBN 1-55670-596-4 (Article: The Humongous Book of Dinosaurs)
- Zhou, Z. (2004). "The origin and early evolution of birds: discoveries, disputes, and perspectives from fossil evidence" (PDF). Naturwissenschaften. 91 (10): 455–471. Bibcode:2004NW.....91..455Z. doi:10.1007/s00114-004-0570-4. PMID 15365634.
وصلات خارجية

- عامة
- صور
- The Science and Art of Gregory S. Paul Influential paleontologist's anatomy art and paintings
- Skeletal Drawing Professional restorations of numerous dinosaurs, and discussions of dinosaur anatomy.
- تسجيلات مرئية
- BBC Nature:Watch dinosaurs brought to life and get experts' interpretations with videos from BBC programmes including Walking with Dinosaurs
- BBC Explainer - Dinosaurs - a complete history in 4 minutes, animation
- شعبية
- Dinosaurs & other extinct creatures: From the Natural History Museum, a well illustrated dinosaur directory.
- Dinosaurnews (www.dinosaurnews.org) The dinosaur-related headlines from around the world. Recent news on dinosaurs, including finds and discoveries, and many links.
- Dinosauria From UC Berkeley Museum of Paleontology Detailed information – scroll down for menu.
- LiveScience.com All about dinosaurs, with current featured articles.
- Zoom Dinosaurs (www.enchantedlearning.com) From Enchanted Learning. Kids' site, info pages and stats, theories, history.
- Dinosaur genus list contains data tables on nearly every published Mesozoic dinosaur genus as of January 2011.
- LiveScience.com Giant Dinosaurs Get Downsized by LiveScience, June 21, 2009
- تقنية
- Palaeontologia Electronica From Coquina Press. Online technical journal.
- Dinobase A searchable dinosaur database, from the University of Bristol, with dinosaur lists, classification, pictures, and more.
- DinoData (www.dinodata.org) Technical site, essays, classification, anatomy.
- Thescelosaurus! By Justin Tweet. Includes a cladogram and small essays on each relevant genera and species.
| شبيهات الأركوصورات الرئيسية |  | |
|---|---|---|
| أركوصورات Crurotarsi | ||
| Avemetatarsalia و أركوصورات Ornithodira | ||
| أركوصورات طائرة | ||
- صفحات تستخدم خطا زمنيا
- CS1 maint: PMC format
- CS1 errors: unsupported parameter
- CS1 errors: ISBN
- CS1 errors: generic name
- CS1 maint: unflagged free DOI
- Automatic taxobox cleanup
- Articles containing إنگليزية-language text
- Pages using Lang-xx templates
- Articles with hatnote templates targeting a nonexistent page
- Pages with empty portal template
- مقالات ناطقة
- ديناصورات
- حيوانات منقرضة
- علم المتحجرات
- علم الحيوان العتيق
- أحفورات وصفت في 1842