بكتيريا زرقاء
| بكتيريا زرقاء Temporal range: 2100–0Ma
| |
|---|---|

| |
| Microscope image of Cylindrospermum, a genus of Cyanobacteria | |
| التصنيف العلمي | |
| النطاق: | الجراثيم |
| غير مصنفة: | الطحالب البرية |
| غير مصنف: | مجموعة الطحالب الزرقاء-الطحالب الداكنة |
| الشعبة: | الطحالب الزرقاء Stanier, 1973 |
| الصف: | الطحالب الزرقاء |
| Orders[3] | |
|
اعتبارا من 2014[تحديث] the taxonomy was under revision[1][2] | |
| Synonyms | |
| |

تعرف الطحالب الزرقاء المخضرة Cyanobacteria أيضا باسم الطحالب اللزجة Myxophyta وهي تعتبر أبسط أنواع الطحالب وأقلها رقيا وأقدمها وجوداً على ظهر الأرض ، شوهد بعضها في حفريات قديمة .
السيانوبكتريا نشأت في دهر الأركي، وتكون الاستروماتوليتات.[4]
وهي تعيش في بيئات مختلفة ، ويعيش كثيرا من أفرادها في المياه العذبة والمالحة . يعيش بعضها في التربة الرطبة ، أما على سطحها أو بداخلها لعمق قد يصل متر . تعيش الغالبية العظمى من الطحالب الزرقاء المخضرة معيشة ذاتية التغذية ، والقليل منها يعيش رميا أو متطفلا جزئيا على بعض الأنواع الأخرى من الطحالب . يوجد بعضها في ظروف حرارية قاسية فتشاهد أنواع منها في الينابيع الحارة ويشاهد البعض في درجات حرارية منخفضة ، تعيش بعض أنواعها معيشة تعاونية مع بعض الفطريات مكون آشنات lichens .قالب:كتاب مملكة النبات
التركيب

قد يتكون جسم الطحلب من خلية واحدة ، ولكن غالبا ما تتجمع الخلايا وتلتصق معا في مستعمرات مختلفة الأشكال .
تركيب الخلية بسيط ، فالبروتوبلاست خالي من نواة مميزة وميتوكوندريات وجهاز جولجي وبلاستيدات خضراء ولا توجد شبكة اندوبلازمية .
يتكون البروتوبلاست من جزئين عادة ، الخارجي ملون ويعرف بالكروموبلازم chromoplasm والداخلي غير ملون ويعرف بالجسم المركزي central body ويوجد فيه المادة النووية DNA , RNA في حالة انتشار . يرجع لون الكروموبلازم إلى وجود صبغات محمولة على أغشية منتشرة في السيتوبلازم .وهذه الصبغات هي الصبغات الزرقاء فيكوسيانين phycocyanin وكلوروفيل أو كاروتين carotin وزانثوفيل xanthophylls وقد توجد الصبغة الحمراء فيكواريثرين phycoerythrin .
البروتوبلاست ، عادة ، خالي من الفجوات العصارية ، وقد تتكون هذه الفجوات في الخلايا المسنة . وعدم وجود الفجوات العصارية تعتبر من الأسباب التي تجعل الخلية مقاومة للجفاف والضغوط الاسموزية العالية . وتشاهد في بعض الأنواع فجوات كاذبة pseudovacoules ، يعتقد أنها تحتوي على غازات ناتجة عن عملية التنفس وتساعد على طفو الطحالب .
يوجد الغذاء المخزن في صورة قطرات زيت وحبيبات نشا مميز لهذا النوع من الطحالب ويعرف بنشا الطحالب الزرقاء المخضرة cyanophycean starch وهو قريب الشبه بالنشا الحيواني glycogen . وتوجد مواد مخزنة أخرى ، يكثر وجودها في الخلايا التكاثرية وتختفي تدريجيا أثناء فترات النمو والنشاط .
يحاط بروتوبلاست الخلية بجدار سميك يتكون من جزئين ، الخارجي هيميسليلوزي والداخلي يتكون من الميورين murein مشابه في ذلك البكتيريا الموجبة لصبغة جرام ، يحاط الجدار أيضا بغلاف جيلاتيني بكتيني . الجدار قد يكون شفاف عديم اللون وقد يتلون بألوان مختلفة منها الأصفر والأحمر والبنفسجي والبني . ويرجع لون الطحالب إلى مجموعة الألوان الموجودة في الكروموبلازم والملونة للجدار الخارجي للطحالب . وكذا فإن لنون وشدة الضوء المتساقط على الطحلب يؤثر على لون الطحالب .
أفراد هذه الطحالب ليس لها أسواط ولكنها قد تتحرك حركة انزلاقية على السطوح المبتلة ، ويساعد على ذلك إفرازات الطبقة الجيلاتينية المغلفة لها .
الإنقسام والتكاثر
تنقسم خلايا الطحالب الزرقاء المخضرة بالإنقسام البسيط . والقليل منها تنفصل خلاياه مباشرة عقب الإنقسام ، ولكن في معظم الأنواع نجد أن الخلايا الناتجة عن الإنقسام تبقى متصلة ببعضها مكونة مستعمرات وعلى أساس عدد مستويات الإنقسام يتحدد شكل المستعمرة الناتجة . فإذا تتابع الإنقسام في مستوى واحد فإن المستعمرات الناتجة تكون خيطية ، وأحيانا تتفرع خيوط الطحالب ، وإذا حدث الإنقسام في مستويين متعامدين كانت المستعمرة الناتجة ذات شكل طبقي مسطح ، أما إذا حدث الإنقسام في ثلاثة مستويات كانت المستعمرة الناتجة مكعبة الشكل ، وإذا زاد عدد المستويات عن ثلاثة نتجت مستعمرة غير منتظمة الشكل . وغالبا ماتحاط المستعمرة بطبقة جيلاتينية تنتج من خلاياها .
لاتستمر المستعمرة في النمو إلى مالانهاية . بل إنها تتجزأ بعد فترة من نموها ، ويتوقف ذلك على نوع الغلاف الجيلاتيني ، فإذا كان الغلاف رقيقا سهل الذوبان فإن المستعمرة لاتصل إلى حجم كبير ، وإذا كان الغلاف سميكا فإن المستعمرة تكبر في الحجم كثيرا قبل أن تتجزأ . ويرجع تجزؤ المستعمرة إلى موت بعض الخلايا أو ضعف الإتصال بين بعض الخلايا أو تغذية حيوان على جزء منها . وفي الطحالب الخيطية يوجد عادة خلايا أكبر حجما وأغلظ جدرا من الخلايا العادية ، تعرف بالحويصلات المتباينة heterocysts ، وهي خلايا أقل تلونا من باقي الخلايا . والخلية لها ثقبين جانبيين في منطقتي اتصالها بالخلايا الخضرية المجاورة تمر خلالها بلازمودزمات دقيقة microplasmodesmata ، وتسد هذه الثقوب عند النضج ، وتعتبر الحويصلات المتباينة مراكز لتثبيت الآزوت الجوي . تنفصل الخيوط عند الحويصلات المتبياننية عادة ، لضعف اتصالها بالخلايا المجاورة . خلايا الطحالب الخضرية الموجودة بين حويصلتين متباينتين تعرف بالهرموجونة hormogonium ، وعند انفصال الخيوط عندالحويصلات المتباينة تنفصل الهرموجونات ، التي تستقل عن الخيط الأم وتنمو من جديد .
تكون بعض الطحالب الخيطية جراثيما ساكنة غير متحركة تعرف بالأكينيتات akinetes ، وهي تنتج من بعض خلايا الهرموجونة ، وتكون غنية بمحتواها الغذائي وذات جدر أكثر غلظا من الخلايا الأخرى ومقاومة للظروف البيئية غير الملائمة. تنبت الجراثيم عند تحسن الظروف وتكون خيوطا خضرية . في بعض الأجناس تتكون الجراثيم من انقسام بروتوبلاست خلية خضرية مكونة عديدة من الجراثيم الداخلية endospores الصغيرة الحجم .
التكاثر الجنسي غير معروف بين الطحالب الزرقاء المخضرة .
التصنيف
من أفراد قسم الطحالب الزرقاء المخضرة طحلب نوستوك .
- نوستوك Nostoc :
بعض أنواع هذا الطحلب يعيش طافيا على الماء والبعض يعيش معيشة أرضية . طحلب نوستوك يتكون من خلايا كروية تنتظم في مستعمرة خيطية ، توجد بها خلايا بينية أكبر حجما من باقي الخلايا هي الحويصلات المتباينة التي تفصل الهرموجونات (شكل 7 ب) . يحاط خيط النوستوك عادة بغلاف جيلاتيني . وفي الظروف غير الملائمة تتحول بعض خلايا الهرموجونات وخاصة الملاصقة للحويصلات المتباينة إلى جراثيم ساكنة .
بعض أنواع النوستوك يعيش معيشة تعاونية . والبعض يمكنه تثبيت الأزوت الجوي .
Cyanobacteria (النطق الإنگليزي: /saɪˌænoʊbækˈtɪəriə/; also known as blue-green algae, blue-green bacteria, and Cyanophyta) is a phylum of bacteria that obtain their energy through photosynthesis. The name "cyanobacteria" comes from the color of the bacteria (باليونانية: κυανός (kyanós) = blue).
The ability of cyanobacteria to perform oxygenic photosynthesis is thought to have converted the early reducing atmosphere into an oxidizing one, which dramatically changed the composition of life forms on Earth by stimulating biodiversity and leading to the near-extinction of oxygen-intolerant organisms. According to endosymbiotic theory, chloroplasts in plants and eukaryotic algae have evolved from cyanobacterial ancestors via endosymbiosis.
البيئة

Cyanobacteria can be found in almost every conceivable environment, from oceans to fresh water to bare rock to soil. They can occur as planktonic cells or form phototrophic biofilms in fresh water and marine environments, they occur in damp soil, or even on temporarily moistened rocks in deserts. A few are endosymbionts in lichens, plants, various protists, or sponges and provide energy for the host. Some live in the fur of sloths, providing a form of camouflage.
Aquatic cyanobacteria are probably best known for the extensive and highly visible blooms that can form in both freshwater and the marine environment and can have the appearance of blue-green paint or scum. The association of toxicity with such blooms has frequently led to the closure of recreational waters when blooms are observed. Marine bacteriophages are a significant parasite of unicellular marine cyanobacteria. When they infect cells, they lyse them, releasing more phages into the water.[5]
الخصائص
تثبيت النتروجين

Cyanobacteria include unicellular and colonial species. Colonies may form filaments, sheets or even hollow balls. Some filamentous colonies show the ability to differentiate into several different cell types: vegetative cells, the normal, photosynthetic cells that are formed under favorable growing conditions; akinetes, the climate-resistant spores that may form when environmental conditions become harsh; and thick-walled heterocysts, which contain the enzyme nitrogenase, vital for nitrogen fixation. Heterocysts may also form under the appropriate environmental conditions (anoxic) when fixed nitrogen is scarce. Heterocyst-forming species are specialized for nitrogen fixation and are able to fix nitrogen gas into ammonia (NH
3), nitrites (قالب:No2-) or nitrates (قالب:No3-) which can be absorbed by plants and converted to protein and nucleic acids (atmospheric nitrogen is not bioavailable to plants).
Rice plantations utilize healthy populations of nitrogen-fixing cyanobacteria (Anabaena, as symbiotes of the aquatic fern Azolla) for use as rice paddy fertilizer.[6]
Cyanobacteria are arguably the most successful group of microorganisms on earth. They are the most genetically diverse; they occupy a broad range of habitats across all latitudes, widespread in freshwater, marine and terrestrial ecosystems, and they are found in the most extreme niches such as hot springs, salt works, and hypersaline bays. Photoautotrophic, oxygen-producing cyanobacteria created the conditions in the planet's early atmosphere that directed the evolution of aerobic metabolism and eukarotic photosynthesis. Cyanobacteria fulfill vital ecological functions in the world's oceans, being important contributors to global carbon and nitrogen budgets.
– Stewart and Falconer[7]
البيئة
Many cyanobacteria also form motile filaments, called hormogonia, that travel away from the main biomass to bud and form new colonies elsewhere. The cells in a hormogonium are often thinner than in the vegetative state, and the cells on either end of the motile chain may be tapered. In order to break away from the parent colony, a hormogonium often must tear apart a weaker cell in a filament, called a necridium.
Each individual cell of a cyanobacterium typically has a thick, gelatinous cell wall. They lack flagella, but hormogonia and some species may move about by gliding along surfaces. Many of the multi-cellular filamentous forms of Oscillatoria are capable of a waving motion; the filament oscillates back and forth. In water columns some cyanobacteria float by forming gas vesicles, like in archaea. These vesicles are not organelles as such. They are not bounded by lipid membranes but by a protein sheath.
Some of these organisms contribute significantly to global ecology and the oxygen cycle. The tiny marine cyanobacterium Prochlorococcus was discovered in 1986 and accounts for more than half of the photosynthesis of the open ocean.[8] Many cyanobacteria even display the circadian rhythms that were once thought to exist only in eukaryotic cells (see bacterial circadian rhythms).
التمثيل الضوئي

Cyanobacteria account for 20–30%[citation needed] of Earth's photosynthetic productivity and convert solar energy into biomass-stored chemical energy at the rate of ~450 TW. Cyanobacteria utilize the energy of sunlight to drive photosynthesis, a process where the energy of light is used to split water molecules into oxygen, protons, and electrons. While most of the high-energy electrons derived from water are utilized by the cyanobacterial cells for their own needs, a fraction of these electrons are donated to the external environment via electrogenic activity.[9] Cyanobacterial electrogenic activity is an important microbiological conduit of solar energy into the biosphere.
الأيض والعضيات
Cyanobacteria have an elaborate and highly organized system of internal membranes which function in photosynthesis. Cyanobacteria get their name from the bluish pigment phycocyanin, which they use to capture light for photosynthesis. Photosynthesis in cyanobacteria generally uses water as an electron donor and produces oxygen as a by-product, though some may also use hydrogen sulfide as occurs among other photosynthetic bacteria. Carbon dioxide is reduced to form carbohydrates via the Calvin cycle. In most forms the photosynthetic machinery is embedded into folds of the cell membrane, called thylakoids. The large amounts of oxygen in the atmosphere are considered to have been first created by the activities of ancient cyanobacteria. Due to their ability to fix nitrogen in aerobic conditions they are often found as symbionts with a number of other groups of organisms such as fungi (lichens), corals, pteridophytes (Azolla), angiosperms (Gunnera) etc.
Many cyanobacteria are able to reduce nitrogen and carbon dioxide under aerobic conditions, a fact that may be responsible for their evolutionary and ecological success. The water-oxidizing photosynthesis is accomplished by coupling the activity of photosystem (PS) II and I (Z-scheme). In anaerobic conditions, they are also able to use only PS I — cyclic photophosphorylation — with electron donors other than water (hydrogen sulfide, thiosulphate, or even molecular hydrogen[10]) just like purple photosynthetic bacteria. Furthermore, they share an archaeal property, the ability to reduce elemental sulfur by anaerobic respiration in the dark. Their photosynthetic electron transport shares the same compartment as the components of respiratory electron transport. Their plasma membrane contains only components of the respiratory chain, while the thylakoid membrane hosts both respiratory and photosynthetic electron transport.
Attached to thylakoid membrane, phycobilisomes act as light harvesting antennae for the photosystems. The phycobilisome components (phycobiliproteins) are responsible for the blue-green pigmentation of most cyanobacteria. The variations on this theme are mainly due to carotenoids and phycoerythrins which give the cells the red-brownish coloration. In some cyanobacteria, the color of light influences the composition of phycobilisomes. In green light, the cells accumulate more phycoerythrin, whereas in red light they produce more phycocyanin. Thus the bacteria appear green in red light and red in green light. This process is known as complementary chromatic adaptation and is a way for the cells to maximize the use of available light for photosynthesis.
A few genera, however, lack phycobilisomes and have chlorophyll b instead (Prochloron, Prochlorococcus, Prochlorothrix). These were originally grouped together as the prochlorophytes or chloroxybacteria, but appear to have developed in several different lines of cyanobacteria. For this reason they are now considered as part of the cyanobacterial group.
علاقتها بالبلاستيدات الخضراء
|
and similar) and basal cyanobacteria[11]
Chloroplasts found in eukaryotes (algae and plants) likely evolved from an endosymbiotic relation with cyanobacteria. This endosymbiotic theory is supported by various structural and genetic similarities[12] Primary chloroplasts are found among the "true plants" or green plants – species ranging from sea lettuce to evergreens and flowers which contain chlorophyll b – as well as among the red algae and glaucophytes, marine species which contain phycobilins. It now appears that these chloroplasts probably had a single origin, in an ancestor of the clade called Primoplantae. Other algae likely took their chloroplasts from these forms by secondary endosymbiosis or ingestion.
It was once thought that the mitochondria in eukaryotes also developed from an endosymbiotic relationship with cyanobacteria; however, it is now suspected that this evolutionary event occurred when aerobic bacteria were engulfed by anaerobic host cells. Mitochondria are believed to have originated not from cyanobacteria but from an ancestor of Rickettsia.[citation needed]
علاقتها بتاريخ الأرض
Stromatolites of fossilized oxygen-producing cyanobacteria have been found from 2.8 billion years ago,[13] possibly as old as 3.5 billion years ago.


The biochemical capacity to use water as the source for electrons in photosynthesis evolved once, in a common ancestor of extant cyanobacteria. The geologic record indicates that this transforming event took place early in our planet's history, at least 2450-2320 million years ago (mya), and probably much earlier. Geobiological interpretation of Archean (>2500 mya) sedimentary rocks remains a challenge; available evidence indicates that life existed 3500 mya, but the question of when oxygenic photosynthesis evolved continues to engender debate and research.
A clear paleontological window on cyanobacterial evolution opened about 2000 mya, revealing an already diverse biota of blue-greens. Cyanobacteria remained principal primary producers throughout the Proterozoic (2500-543 mya), in part because the redox structure of the oceans favored photoautotrophs capable of nitrogen fixation.
The most common cyanobacterial structures in the fossil record are the mound-producing stromatolites and related oncolites. Indeed, these fossil colonies are so common that paleobiology, micropaleontology and paleobotany cite the Pre-Cambrian and Cambrian period as an "age of stromatolites" and an "age of algae."
Green algae joined the blue-greens as major primary producers on continental shelves near the end of the Proterozoic, but only with the Mesozoic era (251-65 mya) radiations of dinoflagellates, coccolithophorids, and diatoms did primary production in marine shelf waters take modern form.
Today, the blue-green bacteria remain critical to marine ecosystems as primary producers in oceanic gyres, as agents of biological nitrogen fixation, and—in modified form—as the plastids of marine algae.[14]
التصنيف

Historically, Bacteria were first classified as plants constituting the class Schizomycetes, which along with the Schizophyceae (blue green algae/Cyanobacteria) formed the phylum Schizophyta.[15]then in the phylum Monera in the kingdom Protista by Haeckel in 1866, comprising Protogens, Protamaeba, Vampyrella, Protomonae and Vibrio, but not Nostoc and other cyanobacteria, which were classified with algae[16] later reclassified as the Prokaryotes by Chatton.[17]
The cyanobacteria were traditionally classified by morphology into five sections, referred to by the numerals I-V. The first three – Chroococcales, Pleurocapsales, and Oscillatoriales – are not supported by phylogenetic studies. However, the latter two – Nostocales and Stigonematales – are monophyletic, and make up the heterocystous cyanobacteria. The members of Chroococales are unicellular and usually aggregate in colonies. The classic taxonomic criterion has been the cell morphology and the plane of cell division. In Pleurocapsales, the cells have the ability to form internal spores (baeocytes). The rest of the sections include filamentous species. In Oscillatoriales, the cells are uniseriately arranged and do not form specialized cells (akinetes and heterocysts). In Nostocales and Stigonematales the cells have the ability to develop heterocysts in certain conditions. Stigonematales, unlike Nostocales, includes species with truly branched trichomes. Most taxa included in the phylum or division Cyanobacteria have not yet been validly published under the Bacteriological Code. Except:
- The classes Chroobacteria, Hormogoneae and Gloeobacteria
- The orders Chroococcales, Gloeobacterales, Nostocales, Oscillatoriales, Pleurocapsales and Stigonematales
- The families Prochloraceae and Prochlorotrichaceae
- The genera Halospirulina, Planktothricoides, Prochlorococcus, Prochloron, Prochlorothrix.
التكنولوجيا الحيوية والتطبيقا

The unicellular cyanobacterium Synechocystis sp. PCC6803 was the third prokaryote and first photosynthetic organism whose genome was completely sequenced.[18] It continues to be an important model organism.[19] The smallest genomes have been found in Prochlorococcus spp. (1.7 Mb)[20][21] and the largest in Nostoc punctiforme (9 Mb).[22] Those of Calothrix spp. are estimated at 12–15 Mb,[23] as large as yeast.
Some cyanobacteria are sold as food, notably Aphanizomenon flos-aquae and Arthrospira platensis (Spirulina).[24]
Recent research has suggested the potential application of cyanobacteria to the generation of Clean and Green Energy via converting sunlight into electricity.[مطلوب توضيح] Currently efforts are underway to commercialize algae-based fuels such as diesel, gasoline and jet fuel.[9][25][26]
الأخطار الصحية
Some cyanobacteria produce toxins, called cyanotoxins. These include anatoxin-a, anatoxin-as, aplysiatoxin, cylindrospermopsin, domoic acid, microcystin LR, nodularin R (from Nodularia), or saxitoxin. Cyanobacteria reproduce explosively under certain conditions. This results in algal blooms, which can become harmful to other species if the cyanobacteria involved produce toxins.
These toxins can be neurotoxins, hepatotoxins, cytotoxins, and endotoxins, and can be toxic and dangerous to humans as well as other animals and marine life in general. Several cases of human poisoning have been documented but a lack of knowledge prevents an accurate assessment of the risks.[27][28][29] Recent studies suggest that significant exposure to high levels of some species of cyanobacteria causes amyotrophic lateral sclerosis (ALS). The Lake Mascoma ALS cluster [30] and Gulf War veteran's cluster are two notable examples.[28][29][31]
معرض صور
-
 Cyanobacteria activity turns Coatepeque Caldera lake into a turquoise color
Cyanobacteria activity turns Coatepeque Caldera lake into a turquoise color -
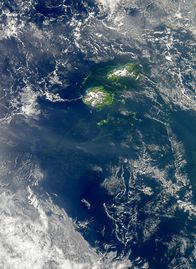 Cyanobacterial bloom near Fiji
Cyanobacterial bloom near Fiji

انظر أيضاً
- Archean Eon
- Bacterial phyla, other major lineages of Bacteria
- Biodiesel
- Cyanobiont
- Endosymbiotic theory
- Geological history of oxygen
- Hypolith
- Microbial mats
- Microalgae
- Phytoplankton
- Proterozoic Eon
المراجع
- ^ Cyanophyceae. Access Science. doi:. Archived from the original. You must specify the date the archive was made using the
|archivedate=parameter. http://www.accessscience.com/content/Cyanophyceae/175300. Retrieved on 21 April 2011. - ^ Oren A (September 2004). "A proposal for further integration of the cyanobacteria under the Bacteriological Code". International Journal of Systematic and Evolutionary Microbiology. 54 (Pt 5): 1895–902. doi:10.1099/ijs.0.03008-0. PMID 15388760.
- ^ Komárek J, Kaštovský J, Mareš J, Johansen JR (2014). "Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach" (PDF). Preslia. 86: 295–335.
- ^ عبد الجليل هويدي، محمد أحمد هيكل (2004). أساسيات الجيولوجيا التاريخية. مكتبة الدار العربية للكتب.
- ^ Photosynthetic viruses keep world's oxygen levels up New Scientist 30 August 2009 by Nora Schultz
- ^ "Azolla-Anabaena as a Biofertilizer for Rice Paddy Fields in the Po Valley, a Temperate Rice Area in Northern Italy". Azolla-Anabaena as a Biofertilizer for Rice Paddy Fields in the Po Valley, a Temperate Rice Area in Northern Italy. International Journal of Agronomy. Retrieved 21 April 2011.
- ^ Stewart I and Falconer IR (2008) "Cyanobacteria and cyanobacterial toxins" Pages 271–296 in Oceans and human health: risks and remedies from the seas, Eds: Walsh PJ, Smith SL and Fleming LE. Academic Press, ISBN 0123725844.
- ^ Steve Nadis, The Cells That Rule the Seas, Scientific American, Nov. 2003 [1]
- ^ أ ب Pisciotta JM, Zou Y, Baskakov IV (2010). Yang, Ching-Hong (ed.). "Light-Dependent Electrogenic Activity of Cyanobacteria". PLoS One. 5 (5): e10821. doi:10.1371/journal.pone.0010821. PMC 2876029. PMID 20520829.
{{cite journal}}: CS1 maint: multiple names: authors list (link) CS1 maint: unflagged free DOI (link) - ^ Champion Hydrogen-Producing Microbe, ScienceDaily, December 15, 2010
- ^ Enrique Flores AH (2008). The Cyanobacteria: Molecular Biology, Genomics and Evolution. Horizon. p. 3. ISBN 1904455158.
- ^ Summarised in Cavalier-Smith (2000), Trends Plant Sci, 5: 174–182.
- ^ Olson JM (2006). "Photosynthesis in the Archean era". Photosyn. Res. 88 (2): 109–17. doi:10.1007/s11120-006-9040-5. PMID 16453059.
- ^ Herrero A and Flores E (editor). (2008). The Cyanobacteria: Molecular Biology, Genomics and Evolution (1st ed.). Caister Academic Press. ISBN 1904455158. ISBN 978-1-904455-15-8 .
{{cite book}}:|author=has generic name (help) - ^ C. Von Nägeli (1857). R. Caspary (ed.). "Bericht über die Verhandlungen der 33. Versammlung deutscher Naturforscher und Aerzte, gehalten in Bonn von 18 bis 24 September 1857". Botanische Zeitung. 15: 749–776.
{{cite journal}}: Unknown parameter|trans_title=ignored (|trans-title=suggested) (help) - ^ Haeckel, Ernst (1867). Generelle Morphologie der Organismen. Reimer, Berlin.
- ^ É. Chatton (1925). "Pansporella perplexa. Réflexions sur la biologie et la phylogénie des protozoaires". Ann. Sci. Nat. Zool. 10-VII: 1–84.
- ^
T. Kaneko; et al. (1996). "Kaneko, T. et al. (1996) Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein-coding regions". DNA Res. 3 (3): 109–136. doi:10.1093/dnares/3.3.109. PMID 8905231.
{{cite journal}}: Explicit use of et al. in:|author=(help) - ^ Tabei Y, Okada K, Tsuzuki M (2007). "Sll1330 controls the expression of glycolytic genes in Synechocystis sp. PCC 6803". Biochem. Biophys. Res. Commun. 355 (4): 1045–50. doi:10.1016/j.bbrc.2007.02.065. PMID 17331473.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^
G. Rocap; et al. (2003). "Genome divergence in two Prochlorococcus ecotypes reflects oceanic niche differentiation". Nature. 424 (6952): 1042–1047. doi:10.1038/nature01947. PMID 12917642.
{{cite journal}}: Explicit use of et al. in:|author=(help) - ^
A. Dufresne; et al. (2003). "Genome sequence of the cyanobacterium Prochlorococcus marinus SS120, a nearly minimal oxyphototrophic genome". Proc. Natl Acad. Sci. USA. 100 (17): 10020–10025. doi:10.1073/pnas.1733211100. PMC 187748. PMID 12917486.
{{cite journal}}: Explicit use of et al. in:|author=(help) - ^
J.C. Meeks; et al. (2001). "An overview of the genome of Nostoc punctiforme, a multicellular, symbiotic cyanobacterium". Photosynth. Res. 70 (1): 85–106
A number of important advances have occurred in cyanobacterial biotechnology in the recent
years. World wide attention is drawn towards cyanobacteria for their possible use in mariculture, food, feed, fuel, fertilizer, colourant, production of various secondary metabolites including vitamins, toxins, enzymes, pharmaceuticals, pharmacological probes and pollution abatement. Only a few cyanobacterial strains (including Spirulina) have been well-characterized or exploited commercially (Thajuddin and Subramanian. Cyanobacterial biodiversity and potential applications in biotechnology. CURRENT SCIENCE, VOL. 89, NO. 1, 10 JULY 2005). doi:10.1023/A:1013840025518. PMID 16228364.
{{cite journal}}: Explicit use of et al. in:|author=(help); line feed character in|pages=at position 7 (help) - ^
M. Herdman; et al. (1979). "Genome size of cyanobacteria". J. Gen. Microbiol. 111: 73–85.
{{cite journal}}: Explicit use of et al. in:|author=(help) - ^ Spolaore P, Joannis-Cassan C, Duran E, Isambert A (2006). "Commercial applications of microalgae". J. Biosci. Bioeng. 101 (2): 87–96. doi:10.1263/jbb.101.87. PMID 16569602.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Blue green bacteria may help generate 'green' electricity, The Hindu, June 21, 2010
- ^ Joule wins key patent for GMO cyanobacteria that create fuels from sunlight, CO2 and water. Biofuels Digest (2010-09-14). Retrieved on 2011-04-06.
- ^ Cyanobacteria, their toxins and health risks
- ^ أ ب Blue-Green Algae (Cyanobacteria) and their Toxins
- ^ أ ب Harmful Bloom in Lake Atitlán, Guatemala from NASA Earth Observatory, retrieved on 9 January 2010.
- ^ Disease Cluster Found at Mascoma Lake , New Hampshire – ALS Research & Treatments – ALS Forum. Als.net. Retrieved on 2011-04-06.
- ^ their toxins and health risks
- النسب
![]() This article incorporates text available under the CC BY 2.5 license.
This article incorporates text available under the CC BY 2.5 license.
للاستزادة
- Cribbs, Gillian (1997). Nature's Superfood: the Blue-Green Algae Revolution (first ed.). Newleaf. ISBN 978-0-7522-0569-4.
{{cite book}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - Savage, Marshall (1994). The Millennial Project: Colonizing the Galaxy in Eight Easy Steps. Little Brown & Co. ISBN 978-0-316-77163-4.
{{cite book}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - Fogg GE, Stewart WD, Fay P, Walsby AE (1973). The Blue-green Algae. London and New York: Academic Press. ISBN 978-0-12-261650-1.
- "Architects of the earth's atmosphere", Introduction to the Cyanobacteria, University of California, Berkeley, 3 February 2006.
- Whitton BA (2002-04-25). "Phylum Cyanophyta (Cyanobacteria)". The Freshwater Algal Flora of the British Isles. Cambridge: Cambridge University Press. ISBN 978-0-521-77051-4.
- Pentecost A, Franke U (2010). "Photosynthesis and calcification of the stromatolitic freshwater cyanobacterium Rivularia". Eur. J. Phycol. 45 (4): 345–353. doi:10.1080/09670262.2010.492914.
- Whitton BA, Potts M, eds. (2000). The Ecology of Cyanobacteria: their Diversity in Time and Space. Springer. ISBN 978-0-7923-4735-4.
- "From Micro-Algae to Blue Oil". ParisTech Review. December 2011. Archived from the original on 17 April 2016. Retrieved 2 March 2012.
وصلات خارجية
- What are Cyanobacteria and What are its Types?
- Webserver for Cyanobacteria Research
- Diving an Antarctic Time Capsule Filled With Primordial Life
| عن العوالق |   | ||||
|---|---|---|---|---|---|
| حسب الحجم | |||||
| Bacterioplankton | |||||
| عوالق نباتية | |||||
| Flagellates | |||||
| هائم حيواني | |||||
| موضوعات متعلقة | Aeroplankton · زراعة · Algal mat · Algal nutrient solutions · Artificial seawater · الاغتذاء الذاتي · Diel vertical migration · Dimethylsulfoniopropionate · Fish diseases and parasites · Heterotroph · Macroalgae · Manta trawl · Marine mucilage · Primary production · ستروماتوليت · Tychoplankton · Zoid · C-MORE · CPR · AusCPR · MOCNESS · SCAR | ||||
| المجموعات |  | |
|---|---|---|
| علم الأحياء الدقيقة | ||
| علم البيئة | ||
| الحياة البحرية | ||
| متعلقة بالبشر | ||
| تقنيات | ||
| أخرى | ||
- CS1 maint: unflagged free DOI
- CS1 errors: generic name
- CS1 errors: unsupported parameter
- CS1 errors: invisible characters
- Short description is different from Wikidata
- Articles with hatnote templates targeting a nonexistent page
- Missing redirects
- مقالات فيها عبارات متقادمة منذ 2014
- جميع المقالات التي فيها عبارات متقادمة
- Articles containing Greek-language text
- Articles with unsourced statements from December 2010
- Articles with unsourced statements from September 2011
- جميع الصفحات التي تحتاج تنظيف
- مقالات بالمعرفة تحتاج توضيح from March 2011
- Articles with imported Creative Commons Attribution 2.5 text
- Cyanobacteria
- Phototrophic bacteria
- تمثيل ضوئي
- جراثيم سلبية الگرام
- Environmental chemistry
- Bacteria phyla