خلية أفقية شبكية
| الخلية الأفقية | |
|---|---|
 مخطط العصبونات الشبكية. | |
| Details | |
| الجهاز | جهاز الإبصار |
| الموقع | الشبكية |
| Identifiers | |
| MeSH | D051248 |
| NeuroLex ID | nifext_40 |
| المصطلحات التشريحية | |
الخلايا الأفقية (إنگليزية: Horizontal cells)، هي الخلايا العصبية المترابطة أفقيًا والتي لها أجسام خلوية في الطبقة النووية الداخلية لشبكية عيون الفقاريات . أنها تساعد على دمج وتنظيم المدخلات من عدة خلايا مستقبلات للضوء . من بين وظائفها ، يُعتقد أن الخلايا الأفقية مسؤولة عن زيادة التباين عبر التثبيط الجانبي والتكيف مع ظروف الإضاءة الساطعة والخافتة. توفر الخلايا الأفقية ردود فعل مثبطة للمستقبلات الضوئية للعصوي والمخروط.[1][2] يُعتقد أنها مهمة لخاصية محيط المركز المناهض للمجالات المستقبلة لأنواع عديدة من الخلايا العقدية الشبكية.[3]
تشمل الخلايا العصبية الأخرى في شبكية العين الخلايا المستقبلة للضوء والخلايا ثنائية القطب وخلايا أماكرين وخلايا العقدة الشبكية.
البُنية
اعتمادًا على الأنواع ، توجد عادةً فئة أو فئتين من الخلايا الأفقية ، مع افتراض نوع ثالث أحيانًا.[1][2]
تمتد الخلايا الأفقية عبر المستقبلات الضوئية وتجمع المدخلات قبل التشابك مع الخلايا المستقبلة للضوء..[1][2] قد تتشابك الخلايا الأفقية أيضًا مع الخلايا ثنائية القطب ، لكن هذا لا يزال غير مؤكد.[1][4]
توجد كثافة أكبر للخلايا الأفقية باتجاه المنطقة المركزية لشبكية العين. في القط ، لوحظ أن الخلايا الأفقية من النوع A لها كثافة 225 خلية / مم 2 بالقرب من مركز الشبكية وكثافة 120 خلية / مم 2 في شبكية العين المحيطية.[5]
من غير المرجح أن تكون الخلايا الأفقية وغيرها من الخلايا العصبية الداخلية في شبكية العين قريبة من الجيران من نفس النوع الفرعي مما قد يحدث بالصدفة ، مما يؤدي إلى "مناطق الاستبعاد" التي تفصل بينها. توفر ترتيبات الفسيفساء آلية لتوزيع كل نوع من الخلايا بالتساوي عبر شبكية العين ، مما يضمن وصول جميع أجزاء المجال البصري إلى مجموعة كاملة من عناصر المعالجة..[5] لبروتينات الغشاء MEGF10 و MEGF11 أدوار حاسمة في تكوين الفسيفساء بواسطة الخلايا الأفقية وخلايا أماكرين في الفئران..[6]
الوظيفة
يتم إزالة الاستقطاب من الخلايا الأفقية عن طريق إطلاق الغلوتامات من المستقبلات الضوئية ، والذي يحدث في غياب الضوء. يؤدي إزالة الاستقطاب عن الخلية الأفقية إلى زيادة استقطاب المستقبلات الضوئية القريبة. على العكس من ذلك ، في الضوء تطلق المستقبلات الضوئية كمية أقل من الغلوتامات ، مما يؤدي إلى فرط استقطاب الخلية الأفقية ، مما يؤدي إلى إزالة استقطاب المستقبلات الضوئية القريبة. وبالتالي ، توفر الخلايا الأفقية ارتجاع سلبي للمستقبلات الضوئية. يقيس الانتشار الجانبي الواسع بشكل معتدل والاقتران بين الخلايا الأفقية بواسطة موصلات الفجوة ، متوسط مستوى الإضاءة الذي يقع على منطقة من سطح الشبكية ، والتي تقوم الخلايا الأفقية بعد ذلك بطرح قيمة متناسبة من إخراج المستقبلات الضوئية للاحتفاظ بإدخال الإشارة إلى الداخل دائرة شبكية العين ضمن نطاق تشغيلها..[1] الخلايا الأفقية هي أيضًا واحدة من مجموعتين من الخلايا العصبية الداخلية المثبطة التي تساهم في محيط خلايا العقدة الشبكية:[2]
إضاءة مركز فرط الاستقطاب الضوئي فرط استقطاب الخلايا الأفقية إزالة الاستقطاب من مستقبلات الضوء المحيط

الآلية الدقيقة التي بواسطتها إزالة استقطاب الخلايا الأفقية مفرطة الاستقطاب للمستقبلات الضوئية غير مؤكدة. على الرغم من أن الخلايا الأفقية تحتوي على حمض الغاما-أمينوبيوتيريك ، فإن الآليات الرئيسية التي من خلالها تمنع الخلايا الأفقية المخاريط ربما لا تتضمن إطلاق حمض الغاما-أمينوبيوتيريك بواسطة الخلايا الأفقية على المخاريط.[4][7][8] من المحتمل أن تساهم آليتان غير مشتركتين أحدهما الآخر في تثبيط الخلية الأفقية لإطلاق الغلوتامات بواسطة المخاريط. تعتمد كلتا الآليتين المفترضتين على البيئة المحمية التي توفرها المشابك اللاصقة التي تصنعها الخلايا الأفقية على المخاريط.[4][9] الآلية المفترضة الأولى هي آلية سريعة للغاية لا تحتوي على تأخير متشابك ، مما يجعلها واحدة من أسرع المشابك المثبطة المعروفة .[4][10][11] الآلية المفترضة الثانية بطيئة نسبيًا مع ثابت زمني يبلغ حوالي 200 مللي ثانية وتعتمد على إطلاق ادينوزين ثلاثي الفوسفات عبر قنوات Pannexin 1 الموجودة على تشعبات الخلية الأفقية التي تغزو الطرف المشبكي المخروطي. يقوم ecto-ATPase NTPDase1 ATP بحلمهة الادينوزين ثلاثي الفوسفات خارج الخلية إلى ادينوزين احادي الفوسفات ومجموعات الفوسفات والبروتونات. تشكل مجموعات الفوسفات والبروتونات محلولًا لدرجة الحموضة مع pKa 7.2 ، والذي يحافظ على درجة الحموضة في الشق المشبكي حمضية نسبيًا. هذا يمنع قنوات Ca 2+ المخروط وبالتالي يقلل من إطلاق الغلوتامات بواسطة المخاريط.[4][11][12][13][14]
يُعتقد أن المناهض المحيطي المركزي للخلايا ثنائية القطب موروث من المخاريط. ومع ذلك ، عندما يتم إجراء التسجيلات من أجزاء من المخروط بعيدة عن النهايات المخروطية التي تتشابك مع الخلايا ثنائية القطب ، يبدو أن المناهض المحيطي المركزي أقل موثوقية في المخاريط منه في الخلايا ثنائية القطب. نظرًا لأن المشابك الفتاكة من الخلايا الأفقية يتم تصنيعها على أطراف مخروطية ، يُعتقد أن المناهض المركزي المحيطي للمخاريط موجود بشكل أكثر موثوقية في النهايات المخروطية..[15]
انظر أيضاً
المصادر
- ^ أ ب ت ث ج Masland, RH (2012). "The neuronal organization of the retina". Neuron. 76 (2): 266–280. doi:10.1016/j.neuron.2012.10.002. PMC 3714606. PMID 23083731.
- ^ أ ب ت ث Demb JB, Singer JH (November 2015). "Functional Circuitry of the Retina". Annu Rev Vis Sci. 1: 263–289. doi:10.1146/annurev-vision-082114-035334. PMC 5749398. PMID 28532365.
- ^ Chaya, Taro; Matsumoto, Akihiro; Sugita, Yuko; Watanabe, Satoshi; Kuwahara, Ryusuke; Tachibana, Masao; Furukawa, Takahisa (2017-07-17). "Versatile functional roles of horizontal cells in the retinal circuit". Scientific Reports (in الإنجليزية). 7 (1): 1–15. doi:10.1038/s41598-017-05543-2. ISSN 2045-2322.
- ^ أ ب ت ث ج Thoreson WB, Mangel SC (September 2012). "Lateral interactions in the outer retina". Prog Retin Eye Res. 31 (5): 407–41. doi:10.1016/j.preteyeres.2012.04.003. PMC 3401171. PMID 22580106.
- ^ أ ب Wässle H, Riemann HJ (March 1978). "The mosaic of nerve cells in the mammalian retina". Proc. R. Soc. Lond. B Biol. Sci. 200 (1141): 441–61. doi:10.1098/rspb.1978.0026. PMID 26058.
- ^ Kay, JN; Chu, MW; Sanes, JR (2012). "MEGF10 and MEGF11 mediate homotypic interactions required for mosaic spacing of retinal neurons". Nature. 483 (7390): 465–9. doi:10.1038/nature10877. PMC 3310952. PMID 22407321.
- ^ Verweij J, Kamermans M, Spekreijse H (December 1996). "Horizontal cells feed back to cones by shifting the cone calcium-current activation range". Vision Res. 36 (24): 3943–53. doi:10.1016/S0042-6989(96)00142-3. PMID 9068848.
- ^ Verweij J, Hornstein EP, Schnapf JL (November 2003). "Surround antagonism in macaque cone photoreceptors". J. Neurosci. 23 (32): 10249–57. doi:10.1523/JNEUROSCI.23-32-10249.2003. PMID 14614083.
- ^ Barnes S (Dec 2003). "Center-surround antagonism mediated by proton signaling at the cone photoreceptor synapse". J Gen Physiol. 122 (6): 653–6. doi:10.1085/jgp.200308947. PMC 2229589. PMID 14610023.
- ^ Kamermans M, Fahrenfort I, Schultz K, Janssen-Bienhold U, Sjoerdsma T, Weiler R (May 2001). "Hemichannel-mediated inhibition in the outer retina". Science. 292 (5519): 1178–80. doi:10.1126/science.1060101. PMID 11349152.
- ^ أ ب Vroman R, Klaassen LJ, Howlett MH, Cenedese V, Klooster J, Sjoerdsma T, Kamermans M (May 2014). "Extracellular ATP hydrolysis inhibits synaptic transmission by increasing ph buffering in the synaptic cleft". PLoS Biol. 12 (5): e1001864. doi:10.1371/journal.pbio.1001864. PMC 4028192. PMID 24844296.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Hirasawa H, Kaneko A (December 2003). "pH changes in the invaginating synaptic cleft mediate feedback from horizontal cells to cone photoreceptors by modulating Ca2+ channels". J. Gen. Physiol. 122 (6): 657–71. doi:10.1085/jgp.200308863. PMC 2229595. PMID 14610018.
- ^ Vessey JP, Stratis AK, Daniels BA, Da Silva N, Jonz MG, Lalonde MR, Baldridge WH, Barnes S (April 2005). "Proton-mediated feedback inhibition of presynaptic calcium channels at the cone photoreceptor synapse". J. Neurosci. 25 (16): 4108–17. doi:10.1523/JNEUROSCI.5253-04.2005. PMID 15843613.
- ^ Davenport CM, Detwiler PB, Dacey DM (January 2008). "Effects of pH buffering on horizontal and ganglion cell light responses in primate retina: evidence for the proton hypothesis of surround formation". J. Neurosci. 28 (2): 456–64. doi:10.1523/JNEUROSCI.2735-07.2008. PMC 3057190. PMID 18184788.
- ^ Byzov AL, Shura-Bura TM (1986). "Electrical feedback mechanism in the processing of signals in the outer plexiform layer of the retina". Vision Res. 26 (1): 33–44. doi:10.1016/0042-6989(86)90069-6. PMID 3012877.
المراجع
- Masland, RH (2012). "The neuronal organization of the retina". Neuron. 76 (2): 266–280. doi:10.1016/j.neuron.2012.10.002. PMC 3714606. PMID 23083731.
وصلات خارجية
تشريح مقلة العين البشرية | |||||||
|---|---|---|---|---|---|---|---|
| الغلالة الليفية (الخارجية) |
| 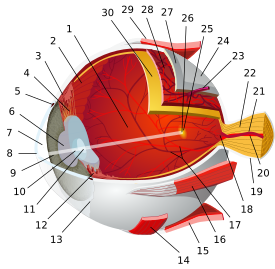  | |||||
| العنبية/ سترة الأوعية الدموية (الوسطى) |
| ||||||
| الشبكية (الداخلية) |
| ||||||
| المناطق التشريحية بالعين |
| ||||||
| غيرهم | |||||||