خلايا الشبكية العقدية الحساسة الجوهرية
.png&filetimestamp=20210331150436&)
خلايا الشبكية العقدية السحاسة الجوهرية (إنگليزية: Intrinsically photosensitive retinal ganglion cells، اختصاراً ipRGCs، تسمى أيضاً خلايا الشبكية العقدية الحساسة للضوء إنگليزية: photosensitive retinal ganglion cells، اختصاراً pRGC، أو الخلايا العقدية الشبكية المحتوية على الميلانوپسين إنگليزية: melanopsin-containing retinal ganglion cells، اختصاراً mRGCs)، هي نوع من الخلايا العصبية في شبكية عين الثدييات . لوحظ وجود الخلايا العقدية الشبكية الحساسة للضوء لأول مرة في عام 1923 عندما كانت الفئران التي ليس لها خلايا عصوية ولا تزال تستجيب لمنبه ضوئي من خلال انقباض حدقة العين ، مما يشير إلى أن العصي والمخاريط ليست الخلايا العصبية الوحيدة الحساسة للضوء في شبكية العين. لم يبدأ التقدم في البحث على هذه الخلايا حتى ثمانينيات القرن الماضي. أظهرت الأبحاث الحديثة أن هذه الخلايا العقدية الشبكية ، على عكس الخلايا العقدية الشبكية الأخرى ، حساسة في جوهرها للضوء بسبب وجود الميلانوبسين ، وهو بروتين حساس للضوء. ولذلك فهي تشكل الطبقة الثالثة من المستقبلات الضوئية، بالإضافة إلى العصي و الخلايا المخروطية.[1]
نظرة عامة

.png&filetimestamp=20210331150434&)
مقارنة بالعصويات والمخاريط ، تستجيب الخلايا العقدية الشبكية الحساسة للضوء ببطء أكبر وتشير إلى وجود الضوء على المدى الطويل.[3] وهي تمثل مجموعة فرعية صغيرة جدًا (حوالي 1٪) من الخلايا العقدية للشبكية.[4] أدوارهم الوظيفية لا تشكل صورة وتختلف جوهريًا عن تلك الخاصة برؤية النمط ؛ أنها توفر تمثيلًا ثابتًا لشدة الإضاءة المحيطة. لديهم ثلاث وظائف أساسية على الأقل:
- يلعبون دورًا رئيسيًا في مزامنة نظم يوماوي مع دورة الضوء / الظلام على مدار 24 ساعة ، مما يوفر معلومات طول اليوم وطول الليل بشكل أساسي. يرسلون معلومات ضوئية عبر السبيل الشبكي المهادي (RHT) مباشرة إلى منظم ضربات القلب اليوماوي في الدماغ ، نواة فوق التصالبة في منطقة ما تحت المهاد .تتطابق الخصائص الفسيولوجية لهذه الخلايا العقدية مع الخصائص المعروفة لآلية تنشيط الضوء اليومي (التزامن) التي تنظم إيقاعات الساعة البيولوجية.. بالإضافة إلى ذلك ، يمكن أن تؤثر الخلايا العقدية الشبكية الحساسة للضوء أيضًا على الأنسجة الطرفية مثل تجديد بصيلات الشعر من خلال دائرة العصب الودي- النواة فوق التصالبية [5]
- تعصب الخلايا العقدية الحساسة للضوء أهداف دماغية أخرى ، مثل مركز التحكم في الحدقة ، نواة أمام السقفية الزيتونية في الدماغ المتوسط . أنها تساهم في تنظيم حجم الحدقة والاستجابات السلوكية الأخرى لظروف الإضاءة المحيطة.[6]
- أنها تساهم في التنظيم الضوئي والقمع الضوئي الحاد لإفراز هرمون الميلاتونين.[6]
- في الفئران ، يلعبون دورًا ما في الإدراك البصري الواعي ، بما في ذلك إدراك الحواجز الشبكية المنتظمة ، ومستويات الضوء ، والمعلومات المكانية.[6]
تم عزل الخلايا العقدية المستقبِلة للضوء في البشر ، حيث ، بالإضافة إلى تنظيم إيقاع الساعة البيولوجية ، ثبت أنها تتوسط درجة من التعرف على الضوء في كائنات ليس لها عصويات او مخاريط تعاني من اضطرابات في المستقبلات الضوئية للقضيب والمخروط..[7] أظهر العمل الذي قام به فرحان زيدي وزملاؤه أن الخلايا العقدية المستقبلة للضوء قد يكون لها بعض الوظائف البصرية لدى البشر.
يتم تحفيز الصبغ الضوئي للخلايا العقدية المستقبلة للضوء ، الميلانوبسين ، بالضوء بشكل رئيسي في الجزء الأزرق من الطيف المرئي (ذروة الامتصاص عند ~ 480 نانومتر[8]. وآلية التنبيغ الضوئي ليست مفهومة بشكل كامل في هذه الخلايا، ولكن يبدو من المرجح أن تشبه التي في رابدوميريك اللافقارية المستقبلات الضوئية. بالإضافة إلى الاستجابة المباشرة للضوء ، قد تتلقى هذه الخلايا تأثيرات استثارة ومثبطة من عصويات ومخاريط عن طريق وصلات متشابكة في شبكية العين.
المحاور من هذه العقد العصبية مناطق الدماغ المتعلقة بالتعرف على الأشياء ، بما في ذلك الأكيمة العلوية والنواة الركبية الجانبية الظهرية.[6]
البنية
مستقبل الخلايا العقدية الشبكية الحساسة للضوء

تنبثق هذه الخلايا المستقبلة للضوء عبر شبكية العين وإلى الدماغ. أنها تحتوي على الميلانوبسين الضوئي بكميات متفاوتة على طول غشاء الخلية ، بما في ذلك على المحاور حتى القرص البصري ، الجسم الجسدي ، والتغصنات في الخلية..[1] تحتوي الخلايا العقدية الشبكية الحساسة للضوء على مستقبلات غشائية للناقلات العصبية الغلوتامات والجلايسين و جابا.[9] تستجيب الخلايا العقدية الحساسة للضوء عن طريق إزالة الاستقطاب ، مما يزيد من معدل إثارة النبضات العصبية ، وهو عكس ذلك الموجود في الخلايا الأخرى المستقبلة للضوء ، والتي تستقطب بشكل مفرط استجابة للضوء.[10]
وتشير نتائج الدراسات على الفئران أن محاور من الخلايا العقدية الشبكية الحساسة للضوء هي عديم الميالين ..[1]
الملانوپسين
على عكس أصباغ مستقبلات الضوء الأخرى ، فإن الميلانوبسين لديه القدرة على العمل كصباغ ضوئي متحمس وكإيزوميراز ضوئي. بدلاً من طلب خلايا إضافية للعودة بين الشكلين الإسفيني ، من كل عبر إلى ريتينال-11 قبل أن تخضع لعملية تنبيغ ضوئي أخرى ، مثل مخاريط مستقبلات الضوء ، التي تعتمد على خلايا مولر وخلايا ظهارة الشبكية الصباغية لهذا التحويل ، الميلانوبسين قادر على تحويل جميع الخلايا عبر الشبكية إلى 11-ريتينال- شبكية عند تحفيزها بالضوء دون مساعدة من خلايا إضافية.[9] يختلف الشكلان الإسويان للميلانوبسين في حساسيتهما الطيفية ، لأن الشكل الإسوي الريتينال -11 هو أكثر استجابة لأطوال موجية أقصر من الضوء ، في حين أن الشكل الإسوي الشامل يكون أكثر استجابة لأطوال موجية أطول من الضوء..[11]
مدخلات ومخرجات متشابكة

المدخلات
تعد الخلايا العقدية الشبكية الحساسة للضوء قبل وبعد المشبكي لخلايا أماكرين دوباميني (خلايا DA) عبر المشابك المتبادلة ، حيث ترسل الخلايا العقدية الشبكية الحساسة للضوء إشارات مثيرة إلى خلايا دوبامينية ، وترسل خلايا دوبامينية إشارات مثبطة إلى الخلايا العقدية الشبكية الحساسة للضوء. يتم التوسط في هذه الإشارات المثبطة من خلال GABA ، والتي يتم إطلاقها بشكل مشترك من خلايا DA جنبًا إلى جنب مع الدوبامين . الدوبامين له وظائف في عملية التكيف مع الضوء عن طريق تنظيم نسخ الميلانوبسين في أماكرين وبالتالي زيادة حساسية مستقبلات الضوء.[1] بالتوازي مع تثبيط خلايا أماكرين دوباميني ، فإن خلايا أماكرين التي تطلق السوماتوستاتين ، والتي تثبط نفسها بواسطة خلايا أماكرينة دوبامينية ، تمنع الخلايا العقدية الشبكية الحساسة للضوء..[12] تشمل المدخلات المشبكية الأخرى للتغصنات الخلايا العقدية الشبكية الحساسة للضوء الخلايا المخروطية ثنائية القطب وخلايا القضيب ثنائية القطب.[9]
الخوارج
أحد أهداف ما بعد المشبكي للخلايا العقدية الشبكية الحساسة للضوء هو النواة فوق التصالبية (SCN) في منطقة ما تحت المهاد ، والتي تعمل بمثابة الساعة البيولوجية في الكائن الحي. تطلق الخلايا العقدية الشبكية الحساسة للضوء كلاً من بروتين تنشيط إنزيم ادينيل سيكليز (PACAP) والغلوتامات على النواة فوق التصالبية عبر اتصال أحادي المشبك يسمى السبيل الشبكي المهادي (RHT).).[13] للجلوتامات تأثير محفز على الخلايا العصبية للنواة فوق التصالبية ، ويبدو أن PACAP يعزز تأثيرات الغلوتامات في منطقة ما تحت المهاد.[14]
تشمل الأهداف الأخرى لما بعد التشابك للخلايا العقدية الشبكية الحساسة للضوء ما يلي: الوريقة بين النواة الركبية (IGL)، وهي مجموعة من الخلايا العصبية الموجودة في المهاد ، والتي تلعب دورًا في التنشيط اليوماوي ؛ النواة أمام السقفية الزيتونية (OPN) ، وهي مجموعة من الخلايا العصبية في الدماغ المتوسط تتحكم في منعكس الحدقة الضوئية ؛ النواة البطنية الجانبية (VLPO) ، الموجودة في منطقة ما تحت المهاد وهي مركز تحكم للنوم; as well as to[clarify] tلوزة المخ.[1]
الوظيفة
منعكس حدقة العين

.png&filetimestamp=20210331150436){kind=link}
{kind=link}
.png&filetimestamp=20210331150434){kind=link}
{kind=link}
{kind=link}
{kind=link}
باستخدام العديد من المستقبلات الضوئية للفئران ، حدد الباحثون دور الخلايا العقدية الشبكية الحساسة للضوء في كل من الإشارات العابرة والمستمرة لمنعكس حدقة العين (PLR)).[15] حدث منعكس حدقة العين العابر في شدة الضوء الخافتة إلى المعتدلة وهو نتيجة التنبيغ الضوئي الذي يحدث في الخلايا العصوية ، والتي توفر مدخلات متشابكة على الخلايا العقدية الشبكية الحساسة للضوء ، والتي بدورها تنقل المعلومات إلى النواة أمام السقفية الزيتونية في الدماغ المتوسط.[16] الناقل العصبي المشترك في ترحيل المعلومات إلى الدماغ المتوسط من الخلايا العقدية الشبكية الحساسة للضوء في منعكس حدقة العين العابر هو الغلوتامات . في شدة الضوء الأكثر إنارةً ، يحدث منعكس حدقة العين المستمر ، والذي يتضمن كلاً من النقل الضوئي للعصوي الذي يوفر مدخلات لللخلايا العقدية الشبكية الحساسة للضوء ونقل ضوئي اللخلايا العقدية الشبكية الحساسة للضوء نفسها عبر الميلانوبسين. اقترح الباحثون أن دور الميلانوبسين في منعكس حدقة العين المستمر يرجع إلى افتقاره للتكيف مع المنبهات الضوئية على عكس الخلايا العصوية ، التي تظهر التكيف. يتم الحفاظ على منعكس حدقة العين متواصل من ببتيد منشط محلقة الأدينيلات النخامية الإفراج عن الخلايا العقدية الشبكية الحساسة للضوء بطريقة نابضة.[15]
دور محتمل في الرؤية الواعية
سمحت التجارب التي أجريت على البشر الذين ليس لديهم عصويات ومخاريط بدراسة دور محتمل آخر للمستقبلات. في عام 2007 ، تم العثور على دور جديد للخلية العقدية المستقبلة للضوء. أظهر زيدي وزملاؤه أن مستقبلات الخلايا العصبية في شبكية العين لدى البشر تساهم في الرؤية الواعية وكذلك في الوظائف غير المكونة للصورة مثل إيقاعات الساعة البيولوجية والسلوك وردود الفعل الحدقة.[7] نظرًا لأن هذه الخلايا تستجيب في الغالب للضوء الأزرق ، فقد اقترح أن لها دورًا في الرؤية الغلسية[بحاجة لمصدر] وأن النظرية القديمة لشبكية العين المزدوجة البحتة مع رؤية عصوية (مظلمة) ومخروطية (ضوئية) كانت مبسطة. عمل الزيدي وزملاؤه مع الأشخاص الذين ليس لديهم عصويات ولا مخاريط ، وبالتالي فتح الباب أيضًا لأدوار تشكيل الصورة (المرئية) لمستقبل الخلية العقدية للضوء.
تم اكتشاف وجود مسارات متوازية للرؤية: أحدهما كلاسيكي قائم على عصوي ومخروط ينشأ من شبكية العين الخارجية ، والآخر كاشف سطوع بصري بدائي ينشأ من الشبكية الداخلية. يبدو أن الأخير يتم تنشيطه بالضوء قبل الأول.[7] تتغذى المستقبلات الضوئية الكلاسيكية أيضًا في نظام المستقبلات الضوئية الجديد ، وقد يكون ثبات اللون دورًا مهمًا على النحو الذي اقترحه فوستر[بحاجة لمصدر].
لقد اقترح مؤلفو النموذج البشري المجهول الخالي من العصي أن المستقبل يمكن أن يكون مفيدًا في فهم العديد من الأمراض ، بما في ذلك الأسباب الرئيسية للعمى في جميع أنحاء العالم مثل الزرق ، وهو مرض يصيب الخلايا العقدية.
في الثدييات الأخرى ، أثبتت العقدة الحساسة للضوء أن لها دورًا حقيقيًا في الرؤية الواعية. الاختبارات التي أجرتها جينيفر ايكر واخرون. وجد أن الفئران التي تفتقر إلى العصويات والمخاريط كانت قادرة على تعلم السباحة باتجاه سلاسل من القضبان العمودية بدلاً من شاشة رمادية مضيئة بنفس القدر.[6]
ضوء بنفسجي إلى أزرق
تشير معظم الأعمال إلى أن حساسية الذروة الطيفية للمستقبل تتراوح بين 460 و 484 نانومتر. لوكلي وآخرون في عام 2003 [17]أظهر أن الطول الموجي 460 نانومتر (الازرق) من الضوء تكبح الميلاتونين ضعف ما يصل إلى 555 نانومتر ضوء(أخضر) ، حساسية الذروة للنظام البصري الضوئي. في عمل زيدي ولوكلي ومؤلفين مشاركين باستخدام إنسان بلا عصوي او مخروطي ، وجد أن منبه كثافته عالية جدًا 481 نانو متر أدى إلى بعض الإدراك الواعي للضوء ، مما يعني أن بعض الرؤية البدائية قد تحققت..[7]
اكتشاف
في عام 1923 ، لاحظ كلايد إي كيلر أن الحدقات في عيون الفئران العمياء التي رعاها عن طريق الخطأ ما زالوا يستجيبون للضوء.[18] كانت قدرة الفئران التي لا تحتوي على عصوي ، ومخاريط على الاحتفاظ بمنعكس حدقة العين توحي بوجود خلية مستقبلة ضوئية إضافية..[9]
في الثمانينيات من القرن الماضي ، أظهرت الأبحاث التي أُجريت على الفئران التي تعاني من نقص العصويات والمخاريط تنظيم الدوبامين في شبكية العين ، وهو معدل عصبي معروف للتكيف مع الضوء والتحفيز الضوئي.[1]
استمر البحث في عام 1991 ، عندما أظهر راسل ج.فوستر وزملاؤه ، بما في ذلك إجناسيو بروفينسيو ، أن العصي والمخاريط ليست ضرورية للحفيز الضوئي ، أو المحرك البصري لإيقاع الساعة البيولوجية ، ولا لتنظيم إفراز الميلاتونين من الغدة الصنوبرية ، عبر عصويات - ومخاريط الفئران.[19][9] أظهر العمل في وقت لاحق من قبل بروفينشو وزملاؤه أن هذة الاستجابة الضوئية تمت بوساطة صباغ ضوئي ميلانوبسين ، موجودة في طبقة الخلايا العقدية للشبكية.[20]
تم تحديد المستقبلات الضوئية في عام 2002 من قبل سامر حتار وديفيد بيرسون وزملاؤهما ، حيث تبين أنها الميلانوبسين تعبر عن الخلايا العقدية التي تمتلك استجابة ضوئية جوهرية وتم إسقاطها على عدد من مناطق الدماغ المشاركة في الرؤية غير المكونة للصورة..[21][22]
في عام 2005 ، أظهر باندا وميليان وكيو وزملاؤه أن صبغة الميلانوبسين الضوئية هي صبغة التنبيغ الضوئي في الخلايا العقدية.[23][24] أظهر دينيس داسي وزملاؤه في نوع من قرود العالم القديم أن الخلايا العقدية العملاقة التي تعبر عن الميلانوبسين متجهة إلى النواة الركبية الجانبية (LGN)).[25][4] في السابق ، تم عرض فقط الإسقاطات على الدماغ المتوسط (نواة امام السقفية) والوطاء (نوى فوق التصالبية ، SCN). ومع ذلك ، كان الدور البصري للمستقبل لا يزال غير متوقع وغير مثبت.
الأبحاث
الأبحاث في البشر
بذلت محاولات لتعقب المستقبلات لدى البشر ، لكن البشر فرضوا تحديات خاصة وطالبوا بنموذج جديد. على عكس الحيوانات الأخرى ، لا يمكن للباحثين أخلاقيا إحداث فقدان العصي والمخروط سواء وراثيا أو مع المواد الكيميائية من أجل دراسة الخلايا العقدية مباشرة. لسنوات عديدة ، كان يمكن استخلاص الاستنتاجات فقط حول المستقبلات في البشر ، على الرغم من أن هذه كانت ذات صلة في بعض الأحيان.
في عام 2007 ، نشر زيدي وزملاؤه عملهم على البشر الذين ليس لديهم عصويات ومخاريط ، الذين أظهروا أن هؤلاء الأشخاص يحتفظون باستجابات طبيعية للتأثيرات غير المرئية للضوء.[7][26] تم العثور على هوية المستقبلات الضوئية غير العصوية وغير المخروطية في البشر على أنها خلية عقدة في شبكية العين الداخلية كما هو موضح سابقًا في نماذج بلا عصويات ومخاريط في بعض الثدييات الأخرى. تم إجراء العمل باستخدام مرضى يعانون من أمراض نادرة قضت على وظيفة مستقبلات الضوء المخروطية والعصوية التقليدية ولكنها حافظت على وظيفة الخلايا العقدية.[7][26] على الرغم من عدم وجود عصويات أو مخاريط ، استمر المرضى في إظهار التحفيز الضوئي اليومي ، والأنماط السلوكية اليومية ، وقمع الميلاتونين ، وردود فعل الحدقات ، مع حساسية طيفية قصوى للضوء البيئي والتجريبي الذي يتطابق مع صبغ الميلانوبسين الضوئي. يمكن لأدمغتهم أيضًا أن تربط الرؤية بضوء هذا التردد. يسعى الأطباء والعلماء الآن إلى فهم دور المستقبل الجديد في الأمراض التي تصيب الإنسان والعمي.[27] كما تم ربط الخلايا العقدية الشبكية الحساسة للضوء في تفاقم الصداع بالضوء أثناء نوبات الصداع النصفي [28]
انظر أيضاً
المصادر
- ^ أ ب ت ث ج ح Do MT, Yau KW (October 2010). "Intrinsically photosensitive retinal ganglion cells". Physiological Reviews. 90 (4): 1547–81. doi:10.1152/physrev.00013.2010. PMC 4374737. PMID 20959623.
- ^ Blume C, Garbazza C, Spitschan M (2019). "Effects of light on human circadian rhythms, sleep and mood". Somnologie (Berl). 23 (3): 147–156. doi:10.1007/s11818-019-00215-x. PMC 6751071. PMID 31534436.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Wong KY, Dunn FA, Berson DM (December 2005). "Photoreceptor adaptation in intrinsically photosensitive retinal ganglion cells". Neuron. 48 (6): 1001–10. doi:10.1016/j.neuron.2005.11.016. PMID 16364903.
- ^ أ ب Berson DM (June 2003). "Strange vision: ganglion cells as circadian photoreceptors". Trends in Neurosciences. 26 (6): 314–20. doi:10.1016/S0166-2236(03)00130-9. PMID 12798601. S2CID 15149809.
- ^ Fan SM, Chang YT, Chen CL, Wang WH, Pan MK, Chen WP, et al. (July 2018). "External light activates hair follicle stem cells through eyes via an ipRGC-SCN-sympathetic neural pathway". Proceedings of the National Academy of Sciences of the United States of America. 115 (29): E6880–E6889. doi:10.1073/pnas.1719548115. PMC 6055137. PMID 29959210.
- ^ أ ب ت ث ج Ecker JL, Dumitrescu ON, Wong KY, Alam NM, Chen SK, LeGates T, et al. (July 2010). "Melanopsin-expressing retinal ganglion-cell photoreceptors: cellular diversity and role in pattern vision". Neuron. 67 (1): 49–60. doi:10.1016/j.neuron.2010.05.023. PMC 2904318. PMID 20624591.
- ^ أ ب ت ث ج ح Zaidi FH, Hull JT, Peirson SN, Wulff K, Aeschbach D, Gooley JJ, et al. (December 2007). "Short-wavelength light sensitivity of circadian, pupillary, and visual awareness in humans lacking an outer retina". Current Biology. 17 (24): 2122–8. doi:10.1016/j.cub.2007.11.034. PMC 2151130. PMID 18082405.
{{cite journal}}: Unknown parameter|lay-date=ignored (help); Unknown parameter|lay-source=ignored (help); Unknown parameter|lay-url=ignored (help) - ^ Berson DM (August 2007). "Phototransduction in ganglion-cell photoreceptors". Pflugers Archiv. 454 (5): 849–55. doi:10.1007/s00424-007-0242-2. PMID 17351786.
- ^ أ ب ت ث ج Kolb H, Fernandez E, Nelson R (1995-01-01). Kolb H, Fernandez E, Nelson R (eds.). Webvision: The Organization of the Retina and Visual System. University of Utah Health Sciences Center. PMID 21413389.
- ^ Do MT, Kang SH, Xue T, Zhong H, Liao HW, Bergles DE, Yau KW (January 2009). "Photon capture and signalling by melanopsin retinal ganglion cells". Nature. 457 (7227): 281–7. Bibcode:2009Natur.457..281D. doi:10.1038/nature07682. PMC 2794210. PMID 19118382.
- ^ Chellappa SL, Ly JQ, Meyer C, Balteau E, Degueldre C, Luxen A, et al. (April 2014). "Photic memory for executive brain responses". Proceedings of the National Academy of Sciences of the United States of America. 111 (16): 6087–91. Bibcode:2014PNAS..111.6087C. doi:10.1073/pnas.1320005111. PMC 4000819. PMID 24616488.
- ^ Vuong HE, Hardi CN, Barnes S, Brecha NC (December 2015). "Parallel Inhibition of Dopamine Amacrine Cells and Intrinsically Photosensitive Retinal Ganglion Cells in a Non-Image-Forming Visual Circuit of the Mouse Retina". The Journal of Neuroscience. 35 (48): 15955–70. doi:10.1523/jneurosci.3382-15.2015. PMC 4666919. PMID 26631476.
- ^ Colwell CS, Michel S, Itri J, Rodriguez W, Tam J, Lelièvre V, et al. (November 2004). "Selective deficits in the circadian light response in mice lacking PACAP". American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 287 (5): R1194-201. doi:10.1152/ajpregu.00268.2004. PMID 15217792.
- ^ Butcher GQ, Lee B, Cheng HY, Obrietan K (June 2005). "Light stimulates MSK1 activation in the suprachiasmatic nucleus via a PACAP-ERK/MAP kinase-dependent mechanism". The Journal of Neuroscience. 25 (22): 5305–13. doi:10.1523/jneurosci.4361-04.2005. PMC 6724997. PMID 15930378.
- ^ أ ب Keenan WT, Rupp AC, Ross RA, Somasundaram P, Hiriyanna S, Wu Z, et al. (September 2016). "A visual circuit uses complementary mechanisms to support transient and sustained pupil constriction". eLife. 5. doi:10.7554/eLife.15392. PMC 5079752. PMID 27669145.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Gamlin PD, McDougal DH, Pokorny J, Smith VC, Yau KW, Dacey DM (March 2007). "Human and macaque pupil responses driven by melanopsin-containing retinal ganglion cells". Vision Research. 47 (7): 946–54. doi:10.1016/j.visres.2006.12.015. PMC 1945238. PMID 17320141.
- ^ Lockley SW, Brainard GC, Czeisler CA (September 2003). "High sensitivity of the human circadian melatonin rhythm to resetting by short wavelength light". The Journal of Clinical Endocrinology and Metabolism. 88 (9): 4502–5. doi:10.1210/jc.2003-030570. PMID 12970330.
- ^ Keeler CE (October 1928). "Blind Mice". Journal of Experimental Zoology. 51 (4): 495–508. doi:10.1002/jez.1400510404.
- ^ Foster RG, Provencio I, Hudson D, Fiske S, De Grip W, Menaker M (July 1991). "Circadian photoreception in the retinally degenerate mouse (rd/rd)". Journal of Comparative Physiology. A, Sensory, Neural, and Behavioral Physiology. 169 (1): 39–50. doi:10.1007/BF00198171. PMID 1941717. S2CID 1124159.
- ^ Provencio I, Rodriguez IR, Jiang G, Hayes WP, Moreira EF, Rollag MD (January 2000). "A novel human opsin in the inner retina". The Journal of Neuroscience. 20 (2): 600–5. doi:10.1523/jneurosci.20-02-00600.2000. PMC 6772411. PMID 10632589.
- ^ Berson DM, Dunn FA, Takao M (February 2002). "Phototransduction by retinal ganglion cells that set the circadian clock". Science. 295 (5557): 1070–3. Bibcode:2002Sci...295.1070B. doi:10.1126/science.1067262. PMID 11834835. S2CID 30745140.
- ^ Hattar S, Liao HW, Takao M, Berson DM, Yau KW (February 2002). "Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity". Science. 295 (5557): 1065–70. Bibcode:2002Sci...295.1065H. doi:10.1126/science.1069609. PMC 2885915. PMID 11834834.
- ^ Panda S, Nayak SK, Campo B, Walker JR, Hogenesch JB, Jegla T (January 2005). "Illumination of the melanopsin signaling pathway". Science. 307 (5709): 600–4. Bibcode:2005Sci...307..600P. doi:10.1126/science.1105121. PMID 15681390. S2CID 22713904.
- ^ Qiu X, Kumbalasiri T, Carlson SM, Wong KY, Krishna V, Provencio I, Berson DM (February 2005). "Induction of photosensitivity by heterologous expression of melanopsin". Nature. 433 (7027): 745–9. Bibcode:2005Natur.433..745Q. doi:10.1038/nature03345. PMID 15674243. S2CID 24999816.
- ^ Dacey DM, Liao HW, Peterson BB, Robinson FR, Smith VC, Pokorny J, et al. (February 2005). "Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project to the LGN". Nature. 433 (7027): 749–54. Bibcode:2005Natur.433..749D. doi:10.1038/nature03387. PMID 15716953. S2CID 4401722.
- ^ أ ب Coghlan A (2007). "How blind people see sunrise and sunset". New Scientist. 196 (2635–2636): 9. doi:10.1016/S0262-4079(07)63172-8.
- ^ Schor J (2008-04-19). "Blue Light and Melatonin" (web page). Morning Light. Retrieved 2008-05-30.[نشر ذاتي سطري?]
- ^ Noseda R (2010). "A neural mechanism for exacerbation of headache by light". Nature Neuroscience. 13: 239–245. doi:10.1038/nn.2475. PMC 2818758.
وصلات خارجية
- Melanopsin-expressing, Intrinsically Photosensitive Retinal Ganglion Cells, Webvision, University of Utah, US
- ipRGCs Brown University, Rhode Island, US
تشريح مقلة العين البشرية | |||||||
|---|---|---|---|---|---|---|---|
| الغلالة الليفية (الخارجية) |
| 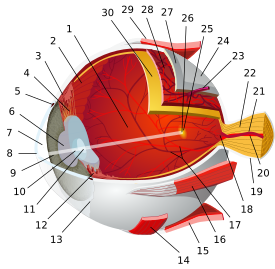  | |||||
| العنبية/ سترة الأوعية الدموية (الوسطى) |
| ||||||
| الشبكية (الداخلية) |
| ||||||
| المناطق التشريحية بالعين |
| ||||||
| غيرهم | |||||||
- CS1 errors: unsupported parameter
- CS1 maint: unflagged free DOI
- All articles with self-published sources
- Articles with self-published sources from June 2014
- Short description is different from Wikidata
- Articles containing إنگليزية-language text
- Pages using Lang-xx templates
- All Wikipedia articles needing clarification
- Wikipedia articles needing clarification from March 2020
- Articles with unsourced statements from November 2010
- Articles with unsourced statements from May 2018
- تشريح العين البشرية
- علم الأنسجة
- خلايا مستقبلة للضوء
- إيقاع يوماوي
- جهاز الابصار
- علم الأعصاب