خلية مظلية
| خلية مظلية | |
|---|---|
 البنية العامة للخلية المظلية. | |
| Details | |
| جزء من | شبكية العين |
| النظام | جهاز الإبصار |
| Identifiers | |
| FMA | 67916 |
| المصطلحات التشريحية | |
الخلية المظلية (إنگليزية: parasol cell، تسمى أحياناً خلية إم (إنگليزية: M cell)[1] أو خلية إم العقدية (إنگليزية: M ganglion cell)،[2] ، هي نوع واحد من الخلايا العقدية الشبكية (RGC) الموجودة في طبقة الخلايا العقدية في شبكية العين . تُسقط هذه الخلايا على الخلايا الخلوية الكبيرة في النواة الركبية الجانبية (LGN) كجزء من المسار الخلوي الكبير في النظام البصري.[3] لديهم أجسام خلوية كبيرة بالإضافة إلى شبكات تغصنية متفرعة واسعة وبالتالي لديهم حقول استقبال كبيرة.[4][3] بالنسبة إلى الخلايا العقدية الشبكية الأخرى ، لديهم سرعات توصيل سريعة..[4] في حين أنهم يظهرون تضادًا واضحًا في الوسط المحيط (يُعرف بالخصم الحيزي) ، إلا أنهم لا يتلقون أي معلومات حول اللون (غياب الخصم اللوني).[3] تساهم الخلايا المظلية بالمعلومات حول حركة الأشياء وعمقها في النظام البصري.[5]
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
الخلايا العقدية المظلية في المسار الخلوي الكبير

الخلايا العقدية المظلية هي الخطوة الأولى في المسار الخلوي الكبير للنظام البصري. إنهم يبرزون من شبكية العين عبر العصب البصري إلى أكثر الطبقتين البطنيتين من النواة الركبية الوحشية ، وهما نواة المهاد ، التي تشغلها الخلايا الخلوية الكبيرة التي تبرز بعد ذلك بشكل أساسي إلى القشرة المخية (V1) ، عادةً إلى الطبقة 4Cα.[6]
في النهاية ، يتم إرسال المعلومات التي تجمعها هذه الخلايا في شبكية العين إلى أجزاء مختلفة من القشرة البصرية ، بما في ذلك القشرة الجدارية الخلفية والمنطقة V5 من خلال التيار الظهري ، والقشرة الصدغية السفلية والمنطقة V4 من خلال التيار البطني[7]
البنية

توجد خلايا العقدة المظلية في شبكية العين ، وتشكل ما يقرب من 10 ٪ من جميع الخلايا العقدية في شبكية العين.[3] لديهم أجسام كبيرة [4][6] مع تغصنات متفرعة واسعة ومتداخلة,[3][8] ومحاور سميكة ، كثيفة الميالين. تسمح هذه الخصائص لخلايا المظلة بتوصيل الإشارات بسرعة كبيرة ، أسرع بكثير من الخلايا القزمة التي تغذي المسار P.[4][6]
تجمع الخلايا العقدية المظلية المعلومات من الحقول المستقبلة الكبيرة,[3][6] التي تحتوي على كل من العصي والمخاريط.[9]على الرغم من المدخلات من المخاريط ، لا تتلقى الخلايا العقدية للمظلة معلومات حول اللون.[3][6] على عكس الخلايا القزمة ، تحتوي الحقول المستقبلة لخلية المظلة على نفس نوع لون المخاريط في كل من مناطقها المركزية والمحيطية. بسبب هذا النقص في النوعية ، لا تستطيع خلايا المظلة التفريق بين أطوال موجات الضوء المختلفة المنعكسة من كائن معين ، وبالتالي يمكنها فقط إرسال معلومات لونية.[10]
توجد تقريبًا نفس كثافة الخلايا العقدية للمظلة في النقرة كما هو الحال في بقية شبكية العين ، وهي خاصية أخرى تميزها عن الخلايا القزمة.[8]
الخلايا المظلية مقابل الخلايا القزمية
تبدأ الخلايا الشبكية القزمية والمظلية المسارين الخلوي الكبير والخلوي الصغير المتوازيان ، على التوالي. بينما تلعب كل من خلايا المظلية والخلايا القزمة دورًا مهمًا في النظام البصري ، فإن تشريحها ومساهماتها الوظيفية تختلف.[3][11][12][13]
| نوع الخلية العقدية الشبكية | الخلية المظلية | الخلية القزمة |
|---|---|---|
| مسار تشارك فيه | مسار خلوي كبير | مسار خلوي صغير |
| حجم جسم الخلية | كبيرة | صغيرة |
| شجرة تغصنية | معقد | افل تعقيدا |
| معدل التوصيل | ~ 1.6 مللي ثانية | ~ 2 مللي ثانية |
| الوظيفة في النظام البصري | "أين" الأشياء ؛ "كيف" لفهم الأشياء | الأشياء "ما" هي وفقا للتفاصيل الدقيقة |
| الحساسية للتردد الحيزي | منخفض | متوسطة الي عالية |
| التردد الزمني | عالي | منخفضة |
| معارضة اللون | غير لوني | الخصم الأحمر والأخضر |
الوظيفة
لا تستطيع الخلايا العقدية للشبكية المظلية توفير معلومات مفصلة أو ملونة بدقة,[4] ولكنها لا تزال توفر معلومات ثابتة وعميقة وحركية مفيدة. تتميز الخلايا العقدية المظلية بدرجة عالية من الضوء / التباين الداكن,[14] وتكون أكثر حساسية عند الترددات الحيزية (الترددات الفراغية) المنخفضة من الترددات الحيزية العالية. نظرًا لمعلومات التباين هذه ، فإن هذه الخلايا جيدة في اكتشاف التغييرات في النصوع ، وبالتالي توفر معلومات مفيدة لأداء مهام البحث المرئي واكتشاف الحواف.[15]
تعتبر الخلايا العقدية للشبكية المظلية مهمة أيضًا لتوفير معلومات حول موقع الأشياء. يمكن لهذه الخلايا اكتشاف اتجاه وموضع الكائنات في الفراغ,[5][12] المعلومات سيتم إرسالها في النهاية من خلال التيار الظهري..[16] هذه المعلومات مفيدة أيضًا في اكتشاف الاختلاف في مواضع الاشياء على شبكية العين لكل عين ، وهي أداة مهمة في إدراك العمق ثنائي العين.[5][17]
تتمتع خلايا المظلة بالقدرة على اكتشاف الترددات الزمنية العالية,[18] وبالتالي يمكنها اكتشاف التغيرات السريعة في موضع الجسم..[6] هذا هو الأساس لاكتشاف الحركة.[5][14][19] تسمح المعلومات المرسلة إلى التلم داخل الفص الجداري (IPS) للقشرة الجدارية الخلفية للمسار الخلوي الكبير بتوجيه الانتباه وتوجيه حركات العين الرمشية لمتابعة الأشياء المتحركة المهمة في المجال البصري.[4][15][19] Iبالإضافة إلى تتبع الكائنات بالعين ، يرسل التلم داخل الفص الجداري معلومات إلى أجزاء من الفص الأمامي والتي تسمح لليدين والذراعين بضبط حركاتهم لفهم الأشياء بشكل صحيح بناءً على حجمها وموضعها وموقعها.[16] قادت هذه القدرة بعض علماء الأعصاب إلى افتراض أن الغرض من المسار الخلوي الكبير ليس اكتشاف المواقع الحيزية(الفراغية) ، ولكن لتوجيه الإجراءات المتعلقة بموضع الأجسام وحركتها.[20]
الأبحاث والتجارب
بينما تُدرس الخلايا العصبية عادةً عن طريق الاستخدام خارج الخلية للأقطاب الكهربائية المعدنية ، تتم دراسة الخلايا العقدية الشبكية على وجه التحديد في المختبر . تسمح هذه الطريقة بتحليل البنية المعقدة والمتشابكة لخلايا المظلة داخل الخلايا. في عام 1941 ، كان بولياك أول عالم يستخدم صبغة جولجي لتحديد خلايا العقد الشبكية. هنا ، تم تحليل التشكل التغصني عن كثب وكشف عن أشجار تغصنية كبيرة. لاحقًا في عام 1986 ، كان كابلان وشابلي أول باحثين يربطون خلايا المظلة بالنظام البصري. تشير تسجيلات جهود S في المحاور المحورية للخلايا العقدية الشبكية في النواة الركبية الوحشية إلى وجود حساسية تباين عالية في الخلايا المنتهية في الطبقة الخلوية الكبيرة من الرئيسيات ؛ يعارضه انخفاض حساسية التباين في الخلايا الموجودة في الطبقة الخلوية الصغيرة..[3]

الرئيسيات وأنظمة النماذج الأخرى
تم استخدام كل من الرئيسيات في العالم القديم والجديد كنظم نموذجية للرؤية البشرية وبالتالي كانت مفيدة في البحث عن خلايا المظلة.[8] العديد من تجارب التوسيم الرجعي باستخدام قرود المكاك ، على سبيل المثال ، ربطت الخلايا العقدية الشبكية المظلية والقزمية مع المسارات الخلوية الكبيرة والخلوية الصغيرة على التوالي. بالإضافة إلى ذلك ، أدت دراسات مماثلة إلى النظريات الكامنة وراء معارضة اللون.[3][8] تدعم الأبحاث التي أجراها داسي (1996) هذه الفكرة حيث عولجت خلايا الشبكية الرئيسية في المختبر بحشوات صبغية. تم العثور على خلايا مظلية للمسار الخلوي الكبير غير لونية.[3] في دراسات أخرى ، ساعدت قرود العالم الجديدة ، مثل قرد القرد (marmosets) ، في الفهم الحالي للتردد الحيزي والزماني للطبقة الخلوية الكبيرة في الواة الركبية الوحشية. باستخدام طريقة صبغة نيسل ، تحتوي الطبقة الكبيرة الخلوية ، بالإضافة إلى الطبقة الخلوية الصغيرة ، على أجسام خلوية أغمق وأكثر كثافة من طبقات الخلايا المحببة ، على سبيل المثال..[11]
تمت دراسة خلايا العقدة الشبكية للقطط ومقارنتها مع تلك الموجودة في النظام البصري لكل من الرئيسيات والبشر. تؤكد الأدلة على الحقول الاستقبالية للقطط أن الحقول الاستقبالية للخلايا المظلية أكبر من تلك الموجودة في الخلايا القزمية بسبب بنيتها الخلوية. من المحتمل أن يوجد نفس الشيء في خلايا الشبكية البشرية مما يسمح بتوطين حيزي أفضل.[3]
الاضطرابات ذات الصلة
ارتبطت الإشارات غير الطبيعية في المسار الخلوي بعسر القراءة والفصام.[21][22]
عسر القراءة
هناك نظرية مفادها أن مشاكل الخلايا العقدية المظلية الغير متطورة قد تساهم في التسبب في عسر القراءة. تساعد معلومات الحركة التي تساهم بها الخلايا العقدية المظلية في نظام الرؤية الدماغ على ضبط العينين بشكل منسق ، وقد تؤدي المشاكل في الحركة الرمشية إلى رؤية ضبابية ومشاكل في القراءة قد يكون هذا التخلف ناتجًا عن عدة عوامل ، بما في ذلك نقص التغذية والطفرات في جين KIAA0319 على الكروموسوم السادس . بالإضافة إلى ذلك ، قد تمنع هجمات المناعة الذاتية التي تقوم بها الأجسام المضادة المضادة للعصبونات نمو خلايا العقدة المظلية المناسبة للعمل بشكل طبيعي ، وهي نظرية تفسر سبب وجود أجهزة المناعة الضعيفة بشكل متكرر لدى الأفراد المصابين بعُسر القراءة.[4]
انظر أيضاً
المصادر
- ^ Brodal, Per (2010). The central nervous system : structure and function (4th ed.). New York: Oxford University Press. pp. 226. ISBN 978-0-19-538115-3.
- ^ Gilbert, Scott F (2004). Purves, Dale (ed.). Neuroscience (3rd ed.). Sunderland, Mass.: Sinauer. p. 274. ISBN 978-0-87893-725-7.
- ^ أ ب ت ث ج ح خ د ذ ر ز س Callaway EM (July 2005). "Structure and function of parallel pathways in the primate early visual system". The Journal of Physiology. 566 (Pt 1): 13–9. doi:10.1113/jphysiol.2005.088047. PMC 1464718. PMID 15905213.
- ^ أ ب ت ث ج ح خ Stein J (2014-01-01). "Dyslexia: the Role of Vision and Visual Attention". Current Developmental Disorders Reports. 1 (4): 267–280. doi:10.1007/s40474-014-0030-6. PMC 4203994. PMID 25346883.
- ^ أ ب ت ث Atkinson, J. (1992-01-01). "Early visual development: differential functioning of parvocellular and magnocellular pathways". Eye. 6 ( Pt 2) (2): 129–135. doi:10.1038/eye.1992.28. PMID 1624034.
- ^ أ ب ت ث ج ح Nassi JJ, Callaway EM (May 2009). "Parallel processing strategies of the primate visual system". Nature Reviews. Neuroscience. 10 (5): 360–72. doi:10.1038/nrn2619. PMC 2771435. PMID 19352403.
- ^ Yabuta, N. H.; Sawatari, A.; Callaway, E. M. (2001-04-13). "Two functional channels from primary visual cortex to dorsal visual cortical areas". Science. 292 (5515): 297–300. doi:10.1126/science.1057916. ISSN 0036-8075. PMID 11303106.
- ^ أ ب ت ث Lee BB (March 1996). "Receptive field structure in the primate retina". Vision Research. 36 (5): 631–44. doi:10.1016/0042-6989(95)00167-0. PMID 8762295.
- ^ Hadjikhani N, Tootell RB (2000-01-01). "Projection of rods and cones within human visual cortex". Human Brain Mapping. 9 (1): 55–63. doi:10.1002/(sici)1097-0193(2000)9:1<55::aid-hbm6>3.0.co;2-u. PMC 6871842. PMID 10643730.
- ^ Sincich, Lawrence C.; Horton, Jonathan C. (2005-01-01). "The circuitry of V1 and V2: integration of color, form, and motion". Annual Review of Neuroscience. 28: 303–326. doi:10.1146/annurev.neuro.28.061604.135731. ISSN 0147-006X. PMID 16022598.
- ^ أ ب Jayakumar J, Dreher B, Vidyasagar TR (May 2013). "Tracking blue cone signals in the primate brain". Clinical & Experimental Optometry. 96 (3): 259–66. doi:10.1111/j.1444-0938.2012.00819.x. hdl:11343/39658. PMID 23186138.
- ^ أ ب Skottun, Bernt C.; Skoyles, John R. (2011-01-01). "On identifying magnocellular and parvocellular responses on the basis of contrast-response functions". Schizophrenia Bulletin. 37 (1): 23–26. doi:10.1093/schbul/sbq114. PMC 3004196. PMID 20929967.
- ^ Skoyles J, Skottun BC (January 2004). "On the prevalence of magnocellular deficits in the visual system of non-dyslexic individuals". Brain and Language. 88 (1): 79–82. doi:10.1016/s0093-934x(03)00162-7. PMID 14698733.
- ^ أ ب Pokorny, Joel (2011-07-07). "Review: steady and pulsed pedestals, the how and why of post-receptoral pathway separation". Journal of Vision. 11 (5): 7. doi:10.1167/11.5.7. PMID 21737512.
- ^ أ ب Cheng, Alicia; Eysel, Ulf T.; Vidyasagar, Trichur R. (2004-10-01). "The role of the magnocellular pathway in serial deployment of visual attention". The European Journal of Neuroscience. 20 (8): 2188–2192. doi:10.1111/j.1460-9568.2004.03675.x. PMID 15450098.
- ^ أ ب Hebart, Martin N.; Hesselmann, Guido (2012-06-13). "What visual information is processed in the human dorsal stream?". The Journal of Neuroscience. 32 (24): 8107–8109. doi:10.1523/JNEUROSCI.1462-12.2012. PMC 6703654. PMID 22699890.
- ^ Poggio, G.F.; Poggio, T. (1984). "The analysis of stereopsis". Annual Review of Neuroscience. 7: 379–412. doi:10.1146/annurev.ne.07.030184.002115. PMID 6370081.
- ^ Anderson, Andrew J.; Jiao, Julie; Bui, Bang V. (2015-09-01). "Efficiently Measuring Magnocellular and Parvocellular Function in Human Clinical Studies". Translational Vision Science & Technology. 4 (5): 1. doi:10.1167/tvst.4.5.1. PMC 4559216. PMID 26346944.
- ^ أ ب Vidyasagar, Trichur R. (2004-01-01). "Neural underpinnings of dyslexia as a disorder of visuo-spatial attention". Clinical & Experimental Optometry. 87 (1): 4–10. doi:10.1111/j.1444-0938.2004.tb03138.x. PMID 14720113.
- ^ Goodale, M.A.; Westwood, D.A. (2004). "An evolving view of duplex vision: separate but interacting cortical pathways for perception and action". Current Opinion in Neurobiology. 14 (2): 203–211. doi:10.1016/j.conb.2004.03.002. PMID 15082326.
- ^ Stein, John (2014-01-01). "Dyslexia: the Role of Vision and Visual Attention". Current Developmental Disorders Reports. 1 (4): 267–280. doi:10.1007/s40474-014-0030-6. ISSN 2196-2987. PMC 4203994. PMID 25346883.
- ^ Bortolon, Catherine; Capdevielle, Delphine; Raffard, Stéphane (2015-06-01). "Face recognition in schizophrenia disorder: A comprehensive review of behavioral, neuroimaging and neurophysiological studies". Neuroscience and Biobehavioral Reviews. 53: 79–107. doi:10.1016/j.neubiorev.2015.03.006. ISSN 1873-7528. PMID 25800172.
تشريح مقلة العين البشرية | |||||||
|---|---|---|---|---|---|---|---|
| الغلالة الليفية (الخارجية) |
| 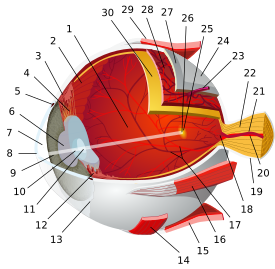  | |||||
| العنبية/ سترة الأوعية الدموية (الوسطى) |
| ||||||
| الشبكية (الداخلية) |
| ||||||
| المناطق التشريحية بالعين |
| ||||||
| غيرهم | |||||||