استشعار النصاب
استشعار النّصاب Quorum sensing، هو نظام تنبيه واستجابة، مرتبط بالكثافة العددية. هناك أنواع كثيرة من الجراثيم تستخدم قدرتَها على استشعار النّصاب لتنظيم التعبير المورّثي تبعا لكثافتها العددية في حيّز مكاني ما، كما أنّ بعض الحشرات الاجتماعية تستخدم نفس الأسلوب لتحديد المكان الذي ستعشّش فيه. وإضافة إلى دور استشعار النّصاب في الأحياء، فإنّ له تطبيقات مفيدة عدّة في الحوسبة وصناعة الروبوتات، حيث يمكن أن يوظَّفَ مبدأُ استشعار النّصاب في عمليات اتخاذ القرار في أي نظام لا مركزي، بشرط أن يشتمل هذا النّظام على:
- وسيلة لتقدير عدد العناصر التي يتفاعل معها كل عنصر من عناصر النّظام.
- استجابة موحَّدة تبديها كافة العناصر إذا تمّ الكشف عن عدد من العناصر يتجاوز حدّا معيّنا.
استشعار النّصاب في الجراثيم
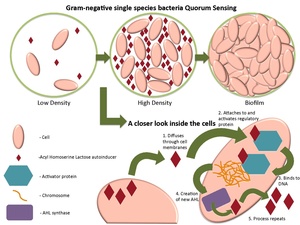
إنّ أشهر الأمثلة على استشعار النّصاب هي تلك التي قدّمتها الدّراسات التي أُجريت على الجراثيم. فالجراثيم تستخدم قدرتها على استشعار النّصاب في تنظيم عدّة سلوكيّات تبعا لكثافتها العدديّة، كتشكيل الأغشية الحيوية الرّقيقة، والفوعة، ومقاومة المضادّات الحيوية.
يمكن أن يتمّ استشعار النّصاب بين خلايا جرثومية من نوع واحد، كما يمكن أن يتمّ بين خلايا تابعة لعدّة أنواع مختلفة.
ينظّم استشعار النّصاب عدّة عمليات مختلفة، تعتمد في أساسها على الكثافة العددية، أو على معدّل انتشار الخلايا في الوسط، حيث تستخدم الجراثيم أنواعا مختلفة من الجزيئات كإشارات تتبادلها فيما بينها، ومن أشهر الجزيئات الإشارية[1]:
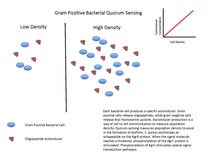
- الببتيدات الصّغيرة (oligopeptides)، وتستخدمها الجراثيم موجبة الگرام.
- اللاكتونات AHL) N-Acyl Homoserine Lactones)، وتستخدمها الجراثيم سالبة الگرام.
- عائلة من المحفّزات الذّاتية (autoinducers (en)) التي تُعرَف بـ المحفّزات الذّاتية-2 (AI-2 (en))، وتستخدمها الجراثيم سالبة الگرام وموجبة الگرام.
آليّات استشعار النصاب
تُنتج الجراثيم التي تستشعر النّصابَ جزيئاتٍ إشاريّةً معيّنة (تسمّى بـ المحفّزات الذّاتية أو الفيرومونات)، وتُفرزها بشكل تلقائي. كذلك فإنّ لهذه الجراثيم مستقبلاتٍ حسّاسة نوعيا لجزيئات الإشارة (المحفّزات). وعندما يرتبط المحفّز بالمستقبِل، يتنشّط نسخَ مورّثات معيّنة، بما فيها تلك المورّثات المسؤولة عن تركيب المحفِّز. ولأنّ هناك احتمالا ضعيفا بأن تتحسّس الجراثيمُ المحفِّزَ الذي أفرزته بنفسها، فلا بدّ من أن تتصادم الخلية بالجزيئات الإشارية التي أفرزتها خلايا أخرى غيرها موجودة معها في نفس الوسط، ليتفعّل النسخ المورّثي.

عندما تكون الخلايا الجرثومية المتجاورة قليلة، وبالتالي تكون متباعدة، فإنّ تباعدَها يخفض من تركيز المحفّز في الوسط المحيط حتى ينعدم تقريبا، فلا تُنتِج إلا القليلَ من المحفّز. ولكن، كلّما ازداد تجمّع الجراثيم، يتخطّى تركيزُ المحفّز العتبةَ، مسببا تركيب المزيد من المحفّز (أي إنّ تركيب المحفّز يخضع لحلقة تغذية راجعة إيجابية)، إلى أن يُصبح المستقبِل مفعّلا تماما، وهذا يزيد نشاطَ مورّثاتٍ خاصّة، مسبّبا بدءَ الخلايا كلّها بالنّسخ في نفس الوقت تقريبا.
إنّ هذا السّلوك المتناسق بين الخلايا الجرثومية، يمكن أن يكون مفيدا في حالات متنوّعة. فمثلا، إنزيم اللوسيفيريز luciferase المتألِّق حيويا، الذي يُنتجه النّوع الجرثومي Vibrio fischeri، لن يكون مرئيا لو أنتجته خلية واحدة فقط. وباستشعار النّصاب، يمكن أن يقتصر إنتاج هذا الإنزيم على الحالات التي يكون فيها التجمّع الخلوي كبيرا، وبذلك تتجنّب خلايا النّوع V. fischeri إهدارَ الطّاقة في إنتاج منتَج لا فائدة منه.
أمثلة
Vibrio fischeri
لوحظ استشعار النّصاب لأوّل مرّة، في النّوع Vibrio fischeri (en)، وهي بكتيريا متألّقة حيويا، تعيش حياةَ تعايش تنافعيّ على حاملات الضّوء (أو الأعضاء المولّدة للضّوء) عند حبّار هاواي قصير الذّيل [2].
عندما تعيش خلايا الجرثوم V. fischeri حياة حرّة (كعوالق مائية، plankton)، يكون تركيز المحفّز الذّاتي منخفضا، وبالتالي، لا تتألّق الخلايا. لكنّها عندما تتجمّع بأعداد كبيرة على الحامل الضّوئي (بمعدّل 1011 خلية/مل تقريبا)، يتحفّز نسخ اللوسيفيريز، ما يؤدّي إلى الإضاءة الحيويّة.
الإشريشيا المعوية
في جراثيم الإشريشيا المعوية سالبة الگرام، يشارك استشعارُ النّصاب في تنظيم الانقسام الخلوي، بواسطة الجزيئات AI-2. فهذه الجراثيم تستخدم الجزيئاتِ AI-2 التي يُنتِجها ويُعالجها الأوبرون lsr، حيث يرمِّز جزءٌ من هذا الأوبيرون ناقلا من النمط ABC (ناقلٌ رابطٌ للأدينوزين ثلاثي الفوسفات، ينقل المواد إلى داخل أو خارج الخلية باستخدام طاقة الأدينوزين ثلاثي الفوسفات)، يُدخِل هذا النّاقلُ جزيئاتِ AI-2 إلى الخلية في أوّل طور النّمو السّاكن أو الثابت (stationary phase)، ثمّ يفسفر إنزيمُ الكيناز LsrK Kinase جزيئاتِ AI-2، وعندها، يمكن أن يبقى الناتج الجديد (AI-2 المفسفَر) داخل الخلية، أو أن يُستخدَم لإيقاف نشاط LsrR، كابح الأوبرون lsr (وبالتالي، يتفعّل الأوبرون).
يُعتقَد أيضا أنّ نسخَ الأوبرون lsr يتثبّط بفوسفات الأسيتون ثنائي الهيدروجين (dihydroxyacetone phosphate، DHAP)، بارتباطه التنافسي مع كابح الأوبرون، LsrR. وقد تبيّن أنّ فوسفات الغليسر ألدهيد (glyceraldehyde 3-phosphate) يثبّط الأوبرون lsr بآليّة التثبيط المتواسَط بـ cAMP-CAPC. وهذا يفسّر عجزَ جراثيم E.coli عن الاحتفاظ بجزيئات AI-2 داخل الخليّة عندما تُنمّى على وسط يحتوي على الغلوكوز (بسبب كبح الاستقلاب الهدمي catabolite repression)، بينما عند تنمّى بشكل طبيعي، تكون جزيئاتُ AI-2 الموجودة في الخليّة نقّآلةً.
ومع أنّ الجراثيم من النّوع إشريشيا معوية أو النّوع سالمونلا كوليرا الخنازير، لا تُنتِج إشارات AHL الشّائعة في البكتيريا موجبة الگرام الأخرى، لكنّها تملك مستقبِلا يكشف وجودَ جزيئات AHL الناتجة عن أنواع جرثومية أخرى، وتُغيّر تعبيرها المورّثي تبعا لوجود تجمّعات "نصابيّة" من البكتيريا سالبة الگرام [3].
سالمونيلا كوليرا الخنازير
ترمّز جراثيم جنس السلمونيلا بروتينا مماثلا للبروتين LuxR، وهو البروتين SdiA، لكنّها لا ترمّز الإنزيمَ AHL synthase (الإنزيم الذي يركّب جزيئاتِ AHL). يكشف SdiA وجودَ الجزيئات AHL التي تنتجها أنواع جرثومية أخرى، بما فيها النّوع Aeromonas hydrophila، والنّوع Hafnia alvei، والنّوع Yersinia enterocolitica [4]. وعندما يتمّ الكشف عن AHL، ينظِّم SdiA عملَ الأوبرون rck الموجود على بلازميد الفوعة في الجنس Salmonella (pefI-srgD-srgA-srgB-rck-srgC)، كما ينظّم عملَ مورّثة واحدة مكتسَبة أفقيا (horizontal acquisition) محمولة على الصّبغي srgE [5][6].
لا تكشف جراثيم السالمونيلا وجودَ جزيئات AHL عندما تمرّ عبر المسالك المعديّة-المعوية لعدّة أنواع حيوانية، وهذا يشير إلى أنّ الأحياء الدّقيقة الموجودة بشكل طبيعي (normal microbiota) في هذه المسالك لا تُنتِج جزيئات AHL، لكنّ البروتين SdiA يتفعَّل عندما تمرّ جراثيم Salmonella عبر المسالك المعوية-المعديّة لسلاحف تستعمرها الجراثيم Aeromonas hydrophila، أو لفئران مخموجة (مُصابة بعدوى) بالجراثيم Yersinia enterocolitica [7][8]. وعلى هذا الأساس، يبدو وكأنّ الجراثيم Salmonella تستخدم البروتين SdiA للكشف عن AHL الذي تُنتجه العوامل الممرضة الأخرى، دون أن تكشف عن البكتيريا الموجودة بشكل طبيعي (الفلورا flora) في القناة الهضمية.
عصيات القيح الأزرق
تستخدم الجراثيم عصيات القيح الأزرق الممرضة الانتهازية قدرتها على استشعار النّصاب لتنسّق تشكيلَ الأغشيّة الحيويّة الرّقيقة، وحركةَ الأسراب، وإنتاجَ كثيرات السكاكر الخارجية، والفوعةَ، والتجمّعَ الخلوي [9]. يمكن أن تنمو هذه الجراثيم في المضيف دون أن تؤذيَه، حتى يصلَ عددها إلى عتبة حديّة، تصبح أعدادُها عندها كافية للتغلّب على الجهاز المناعي للمُضيف. وحينئذ، تبدأ بالهجوم، فتشكّل غشاء حيويا رقيقا، ما يؤدّي إلى مرض المضيف، بينما يشكّل الغشاء الحيوي الرّقيق طبقة واقية تغطّي التجمّعات الجرثومية.
هناك شكل آخر من أشكال التنظيم المورّثي، يمكّن البكتيريا من التكيّف السّريع مع تغيّرات المحيط، وهو يتمّ عبر التأشير البيئي (environmental signaling، وهو التأشير المتبادَل بين الكائن الحي وبيئته)، فقد اكتشفت الدّراسات الحديثة أنّ الحياة بدون أوكسجين (anaerobiosis، أو ما يسمّى بالحياة اللاهوائية) يمكن أن تؤثّر بشكل ملحوظ على الدّارة التنظيمية الرّئيسية لاستشعار النّصاب، وهو ما يؤثّر بدوره على إنتاج الجرثوم لعوامل الفوعة [10].
لقد ثبت تجريبيا أنّ الثّوم والجينسينغ يمنعان استشعار النّصاب في البكتيريا Pseudomonas aeruginosa [11]. ويُؤمل أن يعمل العلاجُ عن طريق التحليل الإنزيمي للجزيئات الإشارية، على منعِ تشكيل أغشية حيوية رقيقة كهذه، أو ربما على إضعافِ الأغشيةَ الحيوية الرّقيقة التي سبق أن تشكّلت حتى. يسمّى تعطيل عملية التأشير بهذه الطّريقة، بـ تثبيط استشعار النّصاب (quorum sensing inhibition).
الراكدة
لقد وُجد حديثا أنّ جراثيم الراكدة، الممرضة الطّارئة، تُبدي قدرةً على استشعار النّصاب [12]. ومن المثير للاهتمام أنّها تبدي نشاطا في تبديد النّصاب (quorum quenching) إضافة إلى ذلك، حيث يمكنها أن تُنتِج جزيئات AHL، كما يمكنها أن تحلّلها أيضا [12].
الأيرومونوس
كانت هذه البكتيريا تُعدّ ممرضة للأسماك، لكنّها برزت مؤخّرا كعامل ممرض للإنسان أيضا، فقد تمّ عزلها من مواقع مخموجة متنوّعة في المرضى (من الصّفراء، والدّم، والسّائل البريتواني، والقيح، والبراز، والبول)، وكلّ العزلات أنتَجت نوعَين أساسيّين من الجزيئات AHL:
- اللاكتون N-butanoyl homoserine lactone (C4-HSL).
- واللاكتون N-hexanoyl homoserine lactone (C6-HSL).
وقد توثّق أنّ النّوع Aeromonas sobria يُنتِج المركّب C6-HSL، إضافة إلى مركّبين أضافّيَّين من الجزيئات AHL، تكون السّلسلةُ الجانبيّة N-acyl فيهما أطولَ منها في المركّب C6 [13].
يرسينيا القولون
إنّ البروتينان YenR و YenI، اللَذان تنتجهما جراثيم يرسينيا القولون التابعة لمجموعة المتقلّبات غاما (gammaproteobacterium)، يشابهان البروتينَين LuxR و LuxI، اللذَين تنتجهما الجراثيم Vibrio fischeri [14][15]. يفعّل YenR تعبيرَ RNA صغير غير مرمِّز للبروتين، اسمه YenS، يثبّط YenS بدوره تعبيرَ البروتين YenI، كما يثبّط إنتاجَ اللاكتون acylhomoserine lactone [16]. يشارك YenR و YenI و YenS في تنظيم السّباحة وحركة الأسراب [15][16].
الجزيئات المشارِكة في استشعار النّصاب
نُشرَت البنية ثلاثية الأبعاد للبروتينات المشارِكة في استشعار النّصاب، لأوّل مرّة، عام 2001، حيث تمّ تحديد البنية البلّوريّة لثلاث بروتيناتٍ موازيةٍ (orthologs) للبروتين LuxS عن طريق تصوير البللورات بالأشعّة السينية [17].
عام 2002، تمّ أيضا تحديد البنية البلّوريّة للمستقبِل LuxP وهو مرتبطٌ بمحفِّزِه AI-2 (وهو من الجزيئات الحيويّة القليلة التي تحتوي على البورون)، في الجراثيم Vibrio haveyi [18].
هناك عدّة أنواع جرثومية تُنتِج الجزيء AI-2، بما فيها جراثيم E. coli المعويّة، التي تُعدّ ممثّلا عن الجراثيم سالبة الگرام. وقد أثبتت أبحاثُ الجينومِ المقارَن وتحليلِ الأصل الشّعبي، التي أُجريت على 138 جينوم من الجراثيم والعتائق archaea وحقيقيّات النّوى، أنّ: " الإنزيم LuxS المطلوب لتركيب AI-2، واسع الانتشار في الجراثيم، بينما يوجد البروتين LuxP، المرتبط بين الغشاءَين الخلويين (periplasmic binding)، في سلالات الجنس Vibrio فقط"، وهذا يدعو إلى استنتاجِ أنّ "الكائنات الحيّة الأخرى إمّا أن تستخدمَ عناصرَ مختلفةً عن نظام النّقل الإشاري AI-2 الذي تستخدمه سلالات الجنس Vibrio، لتستشعر إشارة الجزيء AI-2، أو أنّها لا تملك نظاما كهذا لاستشعار النّصاب على الإطلاق" [19].
تتوفّر قاعدة بيانات لببتيدات استشعار النّصاب تحت الاسم Quorumpeps [20].
هناك بكتيريا معيّنة يمكنها أن تُنتِج إنزيمات تسمّى بإنزيمات اللاكتونيز lactonases، تستهدف الجزيئات AHL وتعطّلها.
تطوّر استشعار النّصاب
تحليل تسلسل مورّثات استشعار النّصاب
إنّ معظمَ نُظُمِ استشعار النّصاب التي تدخل ضمن نموذج "المورّثتَين" (أي إنّ هناك مورّثتين ضروريتين لاستشعار النّصاب، ترمّز إحداهما الإنزيمَ سينتيز synthase الذي يعمل كمحفّز ذاتي، يرتبط مع جزيءٍ مستقبِلٍ ترمّزه المورّثة الأخرى، كنظام البكتيريا Vibrio fischeri الذي سبقَ التعريفُ به)، تُشاهَد في مجموعة المتقلّبات سالبة الگرام. وقد أظهرت المقارنةُ بين الأصلِ الشّعبي للمتقلّبات ـ الذي دُرِس على أساس تسلسلات الـ RNA الريبوزومي ذي معامل التثفيل 16S ـ والأصولِ الشُّعبية لمماثلات (homologs) LuxI أو LuxR أو LuxS، أظهرت تشابها شاملا على درجة عالية. وإجمالا، تبدو مورّثات استشعار النّصاب وكأنّها قد تشعّبت مع تشعّب المتقلّبات، وهذا يشير إلى أنّ نُظم استشعار النّصاب هذه قديمة تماما، وقد ظهرت في وقت مبكّر جدا في سلالة المتقلّبات [21][22].
ورغم وجودِ حالاتٍ تدلّ بوضوح على الانتقال المورّثي الأفقي في الأصول الشّعبية لـ LuxI و LuxR و LuxS، لكنّ مثل هذه الحالات نادرة نسبيا. وهذه النتيجة تتوافق مع الملاحظات التي تشير إلى أنّ مورّثات استشعار النّصاب تنزع إلى التحكّم بتعبير عدد كبير من المورّثات المبعثرة في الصّبغي الجرثومي. ولا يُحتمَل أن يكون اكتسابُ مورّثات الاستشعار قد تمّ حديثا عن طريق الانتقال الأفقي للمورّثات، لأنّ ذلك لا يُتيح لها الزمن الكافي لتتكاملَ إلى هذه الدّرجة. وباعتبار أنّ أغلب الأزواج (المحفّز الذاتي السينتيز/المستقبِل) تقع مترادفة في الجينوم الجرثومي، فمن النادر أن تغيّر قرائنها، ومثل هذه الأزواج يغلب أن تكون قد نشأت معا [22].
إنّ الأصل الشّعبي لمورّثات استشعار النّصاب في المتقلّبات غاما (التي تتضمّن النوعَين Pseudomonas aeroginosa، و Escherichia coli) مثير للاهتمام بشكل خاص، فرغم أنّ مورّثاتها شبيهة وظيفيا بالمورّثتَين LuxI/LuxR (حيث تشكّل المورّثتان LuxI/LuxR زوجَين وظيفيّين: المحفّز الذّاتي السينتيز فيه هو LuxI، والمستقبِل هو LuxR)، لكنّ تسلسلاتها متباعدة إلى حدّ ملفت للنّظر [22]، وهذا يشير إلى أنّ المتقلّبات غاما تتفرّد في طريقة اكتسابها لمورّثات استشعار النّصاب. يمكن أن تكون هذه العائلة من المورّثات المتماثلة قد ظهرت في أسلاف المتقلّبات غاما، رغم أنّ سبب تباعد تسلسلاتها المفرط، مع احتفاظها بالتشابه الوظيفي، ما زال يحتاج تفسيرا. إضافة إلى ذلك، فالأنواع التي تستخدم عدّة نظم غير مترابطة لاستشعار النّصاب، تكون غالبا من المتقلّبات غاما، والدّليل على النّقل الأفقي لمورّثات استشعار النّصاب يكون أوضح ما يمكن في هذا الصّف [21][22].
خلافات في موضوع استشعار النّصاب
بما أنّ مصطلح "استشعار النّصاب" يوحي بأنّ التعاونَ هو الهدفُ من هذه العملية، فقد صار يُنظَر إلى استشعار النّصاب على أنّه الأصل الذي ترتّب عليه نشوءُ الأحياء المحتالة (cheaters) التي تستغلّ التعاونَ بين الأحياء. وقد تم التصدّي لذلك الالتباس باستخدام مفهوم استشعار الانتشار diffusion sensing، الذي شكّل نموذجا بديلا ومكمّلا لمفهوم استشعار النّصاب. مع ذلك، لا يتعرّض هذان المصطلحان لإشكاليّات التأشير، سواءٌ في البيئات البسيطة (خلية واحدة محصورة في حيّز محدّد)، أو المعقّدة (أنواع عديدة تتشارك حيّزا واحدا)، فقد يكون توزّعُ الخلايا مكانيا أهمَّ للاستشعار من الكثافة العددية الخلوية. ولذلك، تمّ طرح نموذج جديد كبديل، يأخذ في الحسبان الإشكاليّتَن معا: الكثافة العددية والحدود المكانية، وسُمّي بـ استشعار الاكتفاء efficiency sensing [23].
يمكن أن يكون أحدُ أسبابِ الخلاف هو أنّ المصطلحات الحاليّة (استشعار النّصاب، واستشعار الكثافة، واستشعار الاكتفاء) كلها فيها إشارة إلى دوافع العملية وفوائدها، وبالتالي، فقد تنطبق على بعض الحالات دون غيرها، ولذلك، ربما يكون إرجاع المصطلح الذي يدل على هذه العملية إلى التحفيز الذاتي، كما وصف هاستينغز (Hastings) وزملاؤه أصلا، هو الحلَّ المعقولَ لهذه الخلافات، حيث أنّ هذا المصطلح لا تُفهَم منه مقاصد العملية أو فوائدها.
المعالجات الطّبية المضادّة لاستشعار النّصاب
إضافة إلى أنّ جزيئات استشعار النّصاب قد تتميّز بفعالية مضادة للميكروبات، يجري الآن التحقق من إمكانية استخدامها ـ وخاصة الببتيدية منها ـ في مجالات علاجية أخرى أيضا، بما فيها مجالات المناعة والأورام. يقوم هذا الافتراض على أساس [24]:
- الدّليلِ الحديث على التأشير المتبادل بين حقيقيّات النّوى وبدائيات النوى، عن طريق جزيئات استشعار النّصاب الإشارية.
- ظاهرة الموت الخلوي المبرمج التي لوحظت في البكتيريا.
- التشابهات الواضحة بين آلية استشعار النّصاب في البكتيريا، والعملية الانتشاريّة التي تنشأ بها الخلايا السّرطانية.
- استهداف مستقبِلات متعدّدة.
- إمكانية التعديل الصّيدلاني على الببتيدات، بحيث ينتج عنه تزايد استهداف المستقبِل.
استشعار النّصاب في العتائقيات
أمثلة
Methanosaeta harundinacea 6Ac
وهو عتائقي مولّد للميتان، يُنتِج مركّبات اللاكتون carboxylated acyl homoserine lactone، التي تُسهّل انتقال النّمو: من النّمو على شكل خليّة قصيرة، إلى النّمو على شكل خيوط [25].
تبديد النّصاب
تبديد النّصاب (quorum quenching) هو منع استشعار النّصاب، عن طريق تشويش التأشير، وقد يتمّ ذلك بتحليل جزيئات التأشير [26][27].
باستخدام الوسط الزّرعي KG، يمكن أن تُعزَل البكتيريا المبدّدة للنّصاب بسهولة من بيئات متنوّعة، بما فيها تلك البكتيريا التي كانت قد اعتُبرت غير قابلة للزرع [27]. ومؤخّرا، تمّ عزل بكتيريا مبدّدة للنّصاب، مدروسة جيّدا، كما دُرست حركيّة تحليلها لمركّب AHL باستخدام الكروماتوغرافيا السائلة سريعة الانحلال (RRLC) [28].
استشعار النّصاب عند الحشرات الاجتماعية
تمثّل مستعمرات الحشرات الاجتماعية نظاما لامركزيا، لأنّه ليس هناك شخص واحد مسؤول عن توجيه المستعمرة أو اتخاذ القرارات عنها. ولقد تبيّن أن عدّة مجموعات من الحشرات الاجتماعية تستخدم استشعار النّصاب بطريقة تشبه اتخاذ القرار الجماعي.
أمثلة
النّمل
تعشّش مستعمرات النّمل من النّوع Temnothorax albipennis في التشقّقات الصّغيرة بين الصّخور، وعندما تتغيّر الصّخور ويُصبح العش مفتوحا، لا بدّ من أن يختار النّمل عشّا جديدا وينتقل إليه بسرعة. في الطّور الأوّل من عملية اتخاذ القرار، يغادر جزء قليل من العاملات العشَّ المدمّر، ويبحث عن صدوع جديدة. وعندما تجد إحدى هؤلاء النّملات الكشّافة مكانا ممكنا للتعشيش، تُقيّم جودةَ الصّدع بناء على عدّة عوامل، منها: حجم الصّدع، وعدد الفتحات (على أساس مستوى الضّوء في الصّدع)، ووجود نمل ميّت أو عدم وجوده [29][30]. تعود العاملة بعدها إلى العشّ المحطّم، حيث ستنتظر فترة قصيرة قبل أن توجِّه العاملاتِ الأخريات إلى العش الذي وجدَتْه، باستخدام عمليّة تُسمّى بالجري المتعاقِب (tandem running (en)). تتناسب فترة الانتظار عكسيا مع جودة الموقع، أي إنّ العاملة التي وجدت موقعا رديئا، ستنتظر فترة أطول من العاملة التي وجدت موقعا جيدا [31]. حين يزور الأفراد الجدد المواقعَ المحتمَلة للعش الجديد، ويقيّمون جودتَها بأنفسهم، يتزايد عدد النملات الزّائرات للصّدوع، ففي هذه المرحلة، قد تزور النّملات مواقع مختلفة للتعشيش، ولكن بسبب اختلاف فترات الانتظار، سيتزايد عدد النّمل في العش الأفضل بمعدّل أعلى من غيره. وأخيرا، سيشعر النّمل في هذا العشّ بأنّ معدّل التقائهم بالنملات الأخريات قد تجاوز حدا معيّنا، وهذا يشير إلى أنّ النّصاب قد اكتمل [32]. وبمجرّد أن يشعر النّمل ببلوغ النّصاب، يعود إلى العش المهدَّم، ويبدأ بحمل الصّغار والملكة والعاملات الأخريات بسرعة إلى العشّ الجديد، كما أنّ الكشّافة التي ما زالت تمارس الجري المتعاقب إلى مواقع أخرى ممكنة للتعشيش، تتوجّه أيضا إلى العش الجديد، وتنتقل المستعمرة كلها. وبالتالي، ورغم أنّ عاملة واحدة لم تكن قد زارت كل المواقع، أو قارنت بين كلّ الخيارات المتاحة، فإنّ استشعار النّصاب يمكّن المستعمرة ككل من اتخاذ قرارات صائبة حول المكان الذي ستنتقل إليه.
نحل العسل
يستخدم نحل العسل Apis mellifera قدرتَه على استشعار النّصاب في اتخاذ قرارات حول موقع العش الجديد أيضا. تتكاثر المستعمرات الكبيرة بعملية تُسمّى بالتبرعم (budding)، حيث تغادر الملكةُ قفيرَ النّحل مع قسم من العاملات، لتشكّل عشّا جديدا في مكان آخر. وبعد أن تغادرَ العش، تشكّل العاملات طردا يتدلى من فرع، أو من أيّة بنية متدلّية، ويستمر هذا الطرد أثناء طور اتخاذ القرار، حتى يتمّ اختيار موقع جديد للعش.
تُشابه عمليةُ استشعار النّصاب في نحل العسل الطريقةَ التي يستخدمها النمل Temnothorax من عدّة نواحٍ، حيث يغادر قسم صغير من العاملات السربَ ليبحث عن موقع للعش الجديد، وتقوم كل عاملة بتقييم جودة التجاويف التي تعثر عليها، ثمّ تعود العاملة إلى الطرد، وتوجّه العاملاتِ الأخريات إلى تجويفها الذي وجدته، عن طريق الرقص الاهتزازي waggle dance (en). وبدلا من استخدام طريقة التأخير في الوقت، فإنّ عدد تكرارات الرّقصة التي تؤدّيها العاملة، هو الذي يدلّ على جودة الموقع، فالعاملات اللاتي وجدن مواقع رديئة للتعشيش، يتوقّفن عن الرّقص أوّلا، وبذلك يمكن توجيههنّ إلى المواقع الأفضل. حالما يستشعر زوّار الموقع الجديد أنهم بلغوا النّصاب (10-20 نحلة عادةً)، فإنّهم يعودون إلى السّرب، ويبدؤون باستخدام طريقة توجيه جديدة، تُدعى طريقة التزمير piping. تسبّب هذه الإشارة الاهتزازيّة تحليقَ السّرب، وتوجّهُه إلى موقع العش الجديد.
في فحص تجريبي، تمكّنت أسراب نحل العسل من اختيار الموقع الأفضل للعش، باستخدام عملية اتخاذ القرار هذه، وأصابت في أربعة اختبارات من أصل خمسة [33][34].
الحوسبة وصناعة الرّوبوتات
يمكن أن يُستخدم استشعار النّصاب لتحسين فعالية الشبكات ذاتية التنظيم، كنظام الرّقابة البيئية SECOAS (المستشعِر التجميعي ذاتي التنظيم، Self-Organizing Collegiate sensor). ففي هذا النّظام، تستشعر العقد الفردية أنّ هناك مجموعة من العقد الأخرى تحمل نفس البيانات، فترشّح المجموعة عقدة واحدة فقط لتبلّغ البيانات، وهذا يوفّر الطّاقة [35].
يمكن أن تستفيد الشبكات اللاسلكية المخصّصة (Ad-hoc) من استشعار النّصاب، بحيث تسمح للنظام بالكشف عن ظروف الشّبكة والاستجابة وفقا لها [36].
يمكن أن يُستخدم استشعار النّصاب أيضا لتنسيق سلوك أسراب الروبوتات المستقلّة، فباستخدام عملية شبيهة بالعملية التي يستخدمها النّمل Temnothorax، يمكن أن تتخذ الروبوتات قرارات سريعة توجّه المجموعة، دون الحاجة إلى قائد يوجّهها [37].
انظر أيضا
المصادر
- ^ Annu. Rev. Microbiol. 55. doi:10.1146/annurev.micro.55.1.165. PMID 11544353.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Journal of Bacteriology. 104 (1). PMC 248216. PMID 5473898 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC248216.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير2=ignored (help); Unknown parameter|الأخير3=ignored (help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول2=ignored (help); Unknown parameter|الأول3=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المسار=ignored (help) - ^ Mol. Microbiol. 52 (4). doi:10.1111/j.1365-2958.2004.04054.x. PMID 15130116.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|التاريخ=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ J. Bacteriol. 183 (19). doi:10.1128/JB.183.19.5733-5742.2001. PMC 95466. PMID 11544237 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC95466.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|التاريخ=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ J. Bacteriol. 180 (5). PMC 107006. PMID 9495757 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC107006.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|التاريخ=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ J. Bacteriol. 185 (4). doi:10.1128/JB.185.4.1357-1366.2003. PMC 142872. PMID 12562806 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC142872.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|التاريخ=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ PLoS ONE. 3 (7). doi:10.1371/journal.pone.0002826. PMC 2475663. PMID 18665275 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2475663.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help); Unknown parameter|المحرر=ignored (help)CS1 maint: unflagged free DOI (link) - ^ J. Bacteriol. 192 (1). doi:10.1128/JB.01139-09. PMC 2798265. PMID 19820103 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2798265.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|التاريخ=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Journal of Bacteriology. 184 (4). doi:10.1128/jb.184.4.1140-1154.2002. ISSN 0021-9193. PMC 134825. PMID 11807075 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC134825.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Cornelis, P. (ed.) (2008). Pseudomonas: Genomics and Molecular Biology. Caister Academic Press.
{{cite book}}:|author=has generic name (help); Cite has empty unknown parameters:|lay-date=,|subscription=,|nopp=,|last-author-amp=,|name-list-format=,|lay-source=,|registration=, and|lay-summary=(help) - ^ Microbiology. 151 (4). doi:10.1099/mic.0.27955-0. PMID 16339933.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ أ ب BMC Microbiology. 11. doi:10.1186/1471-2180-11-51. PMC 3062576. PMID 21385437 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3062576.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلفين المشاركين=ignored (help); Unknown parameter|المسار=ignored (help)CS1 maint: unflagged free DOI (link) - ^ Current Microbiology. 62 (1). doi:10.1007/s00284-010-9689-z. PMID 20544198.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلفين المشاركين=ignored (help); Unknown parameter|المسار=ignored (help) - ^ Molecular microbiology. 17 (2). PMID 7494483.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|التاريخ=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلفين المشاركين=ignored (help) - ^ أ ب Journal of bacteriology. 188 (4). doi:10.1128/JB.188.4.1451-1461.2006. PMC 1367215. PMID 16452428 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1367215.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|التاريخ=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلفين المشاركين=ignored (help) - ^ أ ب Molecular microbiology. 80 (2). doi:10.1111/j.1365-2958.2011.07595.x. PMID 21362062.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|التاريخ=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلفين المشاركين=ignored (help) - ^ Structure. 9 (6). doi:10.1016/S0969-2126(01)00613-X. PMID 11435117.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Nature. 415 (6871). doi:10.1038/415545a. PMID 11823863.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help); Unknown parameter|المسار=ignored (help) - ^ BMC Evol. Biol. 4. doi:10.1186/1471-2148-4-36. PMC 524169. PMID 15456522 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC524169.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help); Unknown parameter|المسار=ignored (help)CS1 maint: unflagged free DOI (link) - ^ Nucleic Acids Research, 2012, 1–5. Doi:10.1093/nar/gks1137
- ^ أ ب Microbiology. 147 (Pt 8). PMID 11496014.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ أ ب ت ث Molecular Biology and Evolution. 21 (5). doi:10.1093/molbev/msh097. PMID 15014168.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Nature Reviews Microbiology. 5 (3). doi:10.1038/nrmicro1600. PMID 17304251.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help); Unknown parameter|المسار=ignored (help) - ^ E. Wynendaele, E. Pauwels, C. Van de Wiele, C. Burvenich, B. De Spiegeleer. The potential role of quorum-sensing peptides in oncology" Medical Hypotheses 78 (2012) 814–817.
- ^ Zhang, G. et al. (2012) Acyl homoserine lactone-based quorum sensing in a methanogenic archaeon. The ISME Journal. advanced online publication
- ^ BMC Microbiology. 11. doi:10.1186/1471-2180-11-51. PMC 3062576. PMID 21385437 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3062576.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلفين المشاركين=ignored (help); Unknown parameter|المسار=ignored (help)CS1 maint: unflagged free DOI (link) - ^ أ ب Journal of Industrial Microbiology & Biotechnology. 36 (2). doi:10.1007/s10295-008-0491-x.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلفين المشاركين=ignored (help); Unknown parameter|المسار=ignored (help) - ^ Antonie van Leeuwenhoek. 98 (3). doi:10.1007/s10482-010-9438-0. PMID 20376561.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلفين المشاركين=ignored (help); Unknown parameter|المسار=ignored (help) - ^ Proceedings of the Royal Society B-Biological Sciences. 273 (1583). doi:10.1098/rspb.2005.3312. PMC 1560019. PMID 16555783 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1560019.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Biology Letters. 1 (2). doi:10.1098/rsbl.2005.0302. PMC 1626204. PMID 17148163 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1626204.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Behavioral Ecology and Sociobiology. 50 (4). doi:10.1007/s002650100377.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Behavioral Ecology. 16 (2). doi:10.1093/beheco/ari020.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ Apidologie. 35 (2). doi:10.1051/apido:2004004.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ American Scientist. 94 (3). doi:10.1511/2006.3.220.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|السنة=ignored (help); Unknown parameter|الصفحات=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المؤلف=ignored (help) - ^ SANPA.
{{cite journal}}: Missing or empty|title=(help); Unknown parameter|الأخير2=ignored (help); Unknown parameter|الأخير=ignored (help); Unknown parameter|الأول2=ignored (help); Unknown parameter|الأول=ignored (help); Unknown parameter|السنة=ignored (help); Unknown parameter|العنوان=ignored (help); Unknown parameter|المسار=ignored (help) - ^ Peysakhov, M.; Regli, W. (2005). "Ant inspired server population management in a service based computing environment" in Swarm Intelligence Symposium, Proceedings 2005 IEEE. Proceedings 2005 IEEE Swarm Intelligence Symposium, 2005. SIS 2005: 357–64. doi:10.1109/SIS.2005.1501643.
- ^ Sahin, E.; Franks, N. (2002). "Measurement of Space: From Ants to Robots". Proceedings of WGW 2002: EPSRC/BBSRC International Workshop
قراءات إضافية
- Dedicated issue of Philosophical Transactions B on quorum sensing. بعض المقالات مجانيّة.
وصلات خارجية
- The Quorum Sensing Website
- Cell-to-Cell Communication in Bacteria
- The SECOAS project—Development of a Self-Organising, Wireless Sensor Network for Environmental Monitoring
- Measurement of Space: From Ants to Robots
- Bonnie Bassler: Discovering bacteria's amazing communication system
- Bonnie Bassler's seminar: "Cell-Cell Communication in Bacteria"

{kind=link}